Российский государственный симфонический оркестр кинематографии, Сергей Скрипка, Дмитрий Илларионов, Борис Андрианов : Московская государственная академическая филармония
Российский государственный симфонический оркестр кинематографии
Коллектив ведет свою историю со времен «великого немого», когда в сентябре 1924 года в московском кинотеатре «Арс» на Тверской место перед экраном занял не пианист-тапер, а оркестр. Музыкальное сопровождение фильмов имело успех у зрителей, и вскоре коллектив, которым руководил композитор и дирижер Давид Блок, стал играть и в других кинозалах. Отныне и навсегда судьба оркестра была связана с кинематографом. В 1938 году оркестр, до этого момента не имевший постоянного состава, был приписан к студии «Союздетфильм». В 1943 году оркестр и хор «Союздетфильма» были выделены в самостоятельную единицу, коллектив получил статус государственного.
Оркестр внес свою лепту в создание лучших фильмов довоенного и военного времени выдающихся режиссеров Сергея Эйзенштейна, Всеволода Пудовкина, Григория Александрова, Григория Козинцева, Ивана Пырьева.
В послевоенные десятилетия оркестр записывает музыку ко множеству фильмов, среди которых «Баллада о солдате» Григория Чухрая с музыкой Михаила Зива, «Война и мир» Сергея Бондарчука с музыкой Вячеслава Овчинникова, «Операция “Ы” и другие приключения Шурика», «Кавказская пленница», «Бриллиантовая рука», «Двенадцать стульев», «Иван Васильевич меняет профессию» Леонида Гайдая с музыкой Александра Зацепина, «Тот самый Мюнхгаузен» Марка Захарова (музыка Алексея Рыбникова), «Мэри Поппинс, до свидания!» Леонида Квинихидзе (музыка Максима Дунаевского), «Место встречи изменить нельзя» Станислава Говорухина (музыка Евгения Геворгяна), «Москва слезам не верит» Владимира Меньшова (музыка Сергея Никитина), «Покровские ворота» Михаила Козакова (музыка Георгия Гараняна) и многие другие.
В 1993 году оркестр возглавил Сергей Скрипка, сумевший сохранить коллектив и поддержать его высокий исполнительский уровень. В числе работ оркестра последних лет – запись музыки Эдуарда Артемьева, Юрия Красавина, Артема Васильева, Анджело Бадаламенти и других композиторов к фильмам и сериалам «Солнечный удар» Никиты Михалкова, «Щелкунчик и Крысиный король», «Белые ночи почтальона Алексея Тряпицына» Андрея Кончаловского, «Ликвидация», «Тихий Дон» и «Жизнь и судьба» Сергея Урсуляка, «Брестская крепость» и «Спитак» Александра Котта, «Достоевский», «Бесы» и «Меморандум Парвуса» Владимира Хотиненко, “Weekend” и «Конец прекрасной эпохи» Станислава Говорухина, «Время первых» Дмитрия Киселёва, «Легенда № 17» и «Экипаж» Николая Лебедева, «Сталинград» Фёдора Бондарчука, «Хождение по мукам» Константина Худякова, «Дух Соноры» (по рок-опере «Звезда и смерть Хоакина Мурьеты») и «Литургия оглашенных» Алексея Рыбникова, «Медный всадник России» Василия Ливанова, «Собибор» Константина Хабенского и многим другим.
С оркестром сотрудничали лучшие отечественные композиторы – Исаак Дунаевский, Сергей Прокофьев, Дмитрий Шостакович, Георгий Свиридов, Эдисон Денисов, Альфред Шнитке, София Губайдулина, Андрей Петров, Родион Щедрин, Андрей Эшпай, Гия Канчели, Эдуард Артемьев, Геннадий Гладков, Владимир Дашкевич, Евгений Дога и другие. В разные годы за его пультом стояли Давид Блок, Александр Гаук, Василий Небольсин, Григорий Гамбург, Николай Аносов, Арий Пазовский, Николай Голованов, Геннадий Рождественский, Евгений Светланов, Марк Эрмлер, Газиз Дугашев, Вероника Дударова, Юрий Силантьев, Арнольд Ройтман, Эмин Хачатурян, Юрий Николаевский, Владимир Васильев, Мартин Нерсесян, Давид Штильман, Константин Кримец, Николай Соколов. С коллективом выступали Давид Ойстрах, Эмиль Гилельс, Иван Козловский, Мстислав Ростропович, Галина Вишневская, Борис Гмыря, Владимир Крайнев, Даниил Шафран, Александр Слободяник, Михаил Плетнёв, Дмитрий Хворостовский.
Оркестр – один из самых мобильных российских коллективов, способных играть в любых больших и малых составах, трансформироваться в эстрадный и джазовый ансамбль, совмещать концертную деятельность со студийной работой. Постоянно участвует в абонементах Московской филармонии, лучшими среди которых стали «Живая музыка экрана» на сцене Концертного зала имени Чайковского и «Кино без границ» в зале «Филармония-2». Свыше двух десятков лет сотрудничал с выдающимся лектором-музыковедом Светланой Виноградовой. Участник музыкальных фестивалей «Декабрьские вечера Святослава Рихтера», «Музыка друзей», «Московская осень», «Славянский базар», Фестиваль российской культуры в Индии, крупных телевизионных концертных проектов, среди которых «Призрак оперы» на Первом канале и «Большая опера» на телеканале «Культура». В 2013 году оркестр удостоен Гранта Правительства России.
Постоянно участвует в абонементах Московской филармонии, лучшими среди которых стали «Живая музыка экрана» на сцене Концертного зала имени Чайковского и «Кино без границ» в зале «Филармония-2». Свыше двух десятков лет сотрудничал с выдающимся лектором-музыковедом Светланой Виноградовой. Участник музыкальных фестивалей «Декабрьские вечера Святослава Рихтера», «Музыка друзей», «Московская осень», «Славянский базар», Фестиваль российской культуры в Индии, крупных телевизионных концертных проектов, среди которых «Призрак оперы» на Первом канале и «Большая опера» на телеканале «Культура». В 2013 году оркестр удостоен Гранта Правительства России.
Сергей Скрипка
Бòльшая часть творческой жизни Сергея Скрипки связана с Российским государственным симфоническим оркестром кинематографии. Он начал сотрудничать с этим коллективом в 1977 году, будучи студентом Московской консерватории в классе профессора Лео Гинзбурга, с 1993 года – художественный руководитель и главный дирижер оркестра. Под руководством Сергея Скрипки Оркестр кинематографии озвучил большинство отечественных кинолент, а также ряд зарубежных фильмов. За это время маэстро обрел уважение коллег, а его творчество стало неотъемлемой частью отечественного кинематографа.
Под руководством Сергея Скрипки Оркестр кинематографии озвучил большинство отечественных кинолент, а также ряд зарубежных фильмов. За это время маэстро обрел уважение коллег, а его творчество стало неотъемлемой частью отечественного кинематографа.
Творческая дружба связывает музыканта с известными мастерами кино. Неоднократно на сцену с маэстро и его оркестром выходили выдающиеся режиссеры Эльдар Рязанов, Никита Михалков, Сергей Соловьёв, Пётр Тодоровский, знаменитые артисты, композиторы, сценаристы. Слушателям запомнились яркие программы, посвященные юбилеям Геннадия Гладкова, Эдуарда Артемьева, Александра Зацепина, Алексея Рыбникова, Владимира Дашкевича, Полада Бюльбюль-оглы, Максима Дунаевского, вечера памяти Тихона Хренникова, Андрея Петрова, Исаака Шварца, Евгения Птичкина, Оскара Фельцмана, Микаэла Таривердиева, Никиты Богословского, Кирилла Молчанова, Георгия Гараняна, Ролана Быкова. Больше десяти лет Сергей Скрипка и оркестр кинематографии осуществляют в Московской филармонии уникальный проект – персональный абонемент «Живая музыка экрана».
Сергей Скрипка сотрудничал со многими российскими коллективами, гастролировал в России и за рубежом. Выступал и записывался с известными солистами – Михаилом Плетнёвым, Дмитрием Хворостовским, Сергеем Судзиловским, Александром Ведерниковым, Любовью Казарновской, Алексеем Любимовым, Андреем Диевым, Родионом Замуруевым, Александром Гиндиным, Алексеем Набиулиным, Алёной Баевой, Гайком Казазяном, Никитой Борисоглебским и другими. С 1975 года руководит Жуковским симфоническим оркестром. В течение 27 лет был профессором кафедры оркестрового дирижирования РАМ имени Гнесиных.
В репертуаре дирижера – произведения разных эпох и жанров: от барокко до новейшей музыки, от высокой классики до лучших образцов эстрады. Впервые в России маэстро исполнил ораторию «Страсти по Марку» Рейнхарда Кайзера, а также осуществил первые записи на CD ряда произведений Глиэра, Мосолова, Шебалина, Денисова. В 2018 году Сергей Скрипка провел мировую премьеру реквиема «Девять шагов к преображению» Эдуарда Артемьева.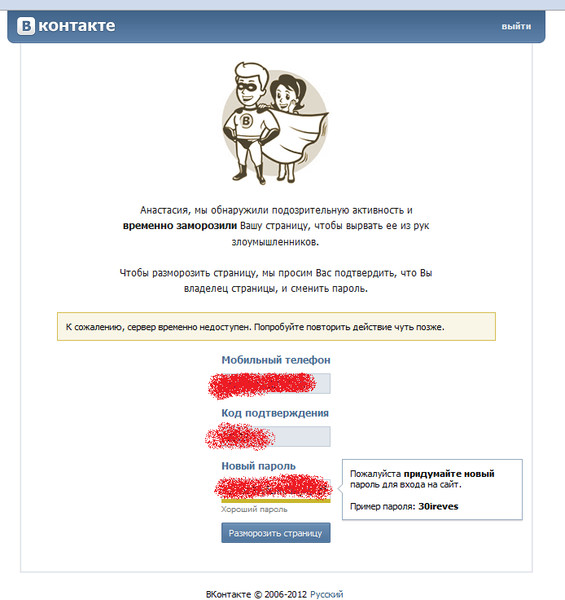
Дмитрий Илларионов
Дмитрий Илларионов родился в 1979 году в Кишинёве. Окончил Академическое музыкальное училище при Московской консерватории (класс Натальи Дмитриевой), Российскую академию музыки имени Гнесиных и ассистентуру-стажировку РАМ (класс профессора Александра Фраучи), затем аспирантуру Высшей школы музыки в Карлсруэ (Германия). В настоящее время – доцент РАМ имени Гнесиных.
Победитель и лауреат 25 международных конкурсов гитаристов в США, Испании, Польше, Италии, Израиле и других странах. Среди них – GFA 2002 (Майами, США), XLII конкурс имени Франсиско Тарреги (Испания, 2008, первый в истории конкурса победитель из России), XXV конкурс имени Андреса Сеговии (Испания, 2009).
Среди них – GFA 2002 (Майами, США), XLII конкурс имени Франсиско Тарреги (Испания, 2008, первый в истории конкурса победитель из России), XXV конкурс имени Андреса Сеговии (Испания, 2009).
Гастролировал в Европе, Азии, Америке. Принимал участие в фестивалях в Шлезвиг-Гольштейне, Москве («Декабрьские вечера Святослава Рихтера», «Возвращение»), имени Пабло Казальса во Франции, Guitar Foundation of America (США), симпозиуме гитары в Изерлоне (Германия). Первый гитарист-солист, принявший участие в фестивале фонда «Орфеум» (Швейцария) и Фестивале камерной музыки в Утрехте (Нидерланды). Среди партнеров гитариста по сцене – Клаудио Шимоне, Лео Брауэр, Денис Мацуев, Патрик Галуа, Эдин Карамазов, Борис Андрианов, квартет Artis и др.
Первый исполнитель ряда сочинений современных автором, в том числе посвященных ему опусов Никиты Кошкина (Concerto Grosso для гитары с оркестром), Анджело Джилардино (концерт «Утренняя звезда» для гитары и виолончели с оркестром памяти Николая Рериха), Сергея Руднева (пьесы для гитары соло), Алексея Рыбникова (сюита «Путешествие во времени» для гитары с оркестром) и др.
Один из основателей Международного фестиваля «Виртуозы гитары» в Москве, руководитель «классического» дня Международного фестиваля «Мир гитары» в Калуге, арт-директор Международного фестиваля гитары в Новосибирске. Член жюри ведущих конкурсов мира: GFA в США, имени Александра Фраучи в Москве, конкурса в Гориции (Италия, 2016, председатель).
Записал сольные диски на фирмах Naxos, Doberman, Daminus. Дуэтный альбом с Борисом Андриановым (Delos) вошел в лонг-лист номинантов премии
Борис Андрианов
Борис Андрианов родился в 1976 году в Москве. Окончил МССМШ имени Гнесиных (класс Веры Бириной), Московскую государственную консерваторию имени П. И. Чайковского (класс профессора Наталии Шаховской) и Высшую школу музыки имени Ханса Айслера в Берлине (класс профессора Давида Герингаса).
И. Чайковского (класс профессора Наталии Шаховской) и Высшую школу музыки имени Ханса Айслера в Берлине (класс профессора Давида Герингаса).
Победитель международных конкурсов в Загребе (имени Антонио Янигро), Йоханнесбурге, Ганновере (Classiсa Nova имени Д. Д. Шостаковича, в дуэте с Алексеем Гориболем), Париже (конкурс Мстислава Ростроповича), Южной Корее (имени Исан Юна). Лауреат XI Международного конкурса имени Чайковского и молодежной премии «Триумф». Удостоен почетного звания «Заслуженный артист Российской Федерации» (2016).
Борис Андрианов выступает с ведущими оркестрами на лучших сценах мира; сотрудничал с известными дирижерами, среди которых Валерий Гергиев, Владимир Федосеев, Юрий Темирканов, Зубин Мета, Юрий Симонов, Павел Коган, Кшиштоф Пендерецкий, Александр Ведерников, Джанандреа Нозеда, Дмитрий Лисс, Роман Кофман, Василий Петренко, Штефан Владар, Давид Герингас, Дмитрий Юровский.
Среди партнеров Андрианова по камерному ансамблю – Юрий Башмет, Менахем Пресслер, Акико Суванаи, Жанин Янсен, Юлиан Рахлин, Сара Макелрави, Итамар Голан, Максим Рысанов, Вадим Холоденко, Филипп Копачевский, Денис Мацуев, Сергей Накаряков и другие.
Участвовал в Шведском Королевском фестивале, фестивалях в Людвигсбурге (Германия), Черво (Италия), Дубровнике (Хорватия), Кольмаре (Франция), Давосе (Швейцария), Иркутске («Звезды на Байкале»), Hiiumaa Homecoming Festival на о. Хийумаа (Эстония) и др. Постоянный участник московского фестиваля камерной музыки «Возвращение».
Организатор и руководитель всероссийского проекта «Поколение звезд» (Премия Правительства РФ в области культуры, 2009). Автор идеи и художественный руководитель международных фестивалей Vivacello и Vivarte в Москве, художественный руководитель фестиваля «Музыкальная экспедиция», проходящего во Владимирской области и других регионах России.
Компакт-диск, записанный Борисом Андриановым с гитаристом Дмитрием Илларионовым на фирме Delos, вошел в лонг-лист номинантов премии «Грэмми» в разделе «камерный ансамбль» (2003). Альбом сонат Шостаковича и Рахманинова, записанный с пианистом Рэмом Урасиным, в сентябре 2007 года был отмечен как лучший камерный диск месяца британским музыкальным журналом Gramophone. В 2015 году на фирме «Мелодия» вышел совместный альбом Бориса Андрианова и Юрия Медяника с записью Сонат для виолы да гамба и клавесина И. С. Баха в транскрипции для виолончели и баяна.
В 2015 году на фирме «Мелодия» вышел совместный альбом Бориса Андрианова и Юрия Медяника с записью Сонат для виолы да гамба и клавесина И. С. Баха в транскрипции для виолончели и баяна.
С 2009 года преподает в Московской консерватории. Играет на инструменте работы Карло Бергонци из Государственной коллекции уникальных музыкальных инструментов.
АНТРОПАТОЛОГИЯ: «Пропагандисты искусственного оплодотворения развлекают нравственное либидо щепотками девственности» (16-й этюдник)
— цикл из книги «Личинки и Лики» (20 этюдников) — написан с 25.09.2001 по 26.09.2001.
«Иисус отвечал: истинно, истинно говорю тебе, если кто не родится от воды и Духа, не может войти в Царствие Божие. Рожденное от плоти есть плоть, а рожденное от Духа есть дух.»
(ОТ ИОАННА СВЯТОЕ БЛАГОВЕСТВОВАНИЕ)
«Ганоидных рыб ставят между селахиями и костистыми; последние в настоящее время представляют группу, по числу форм значительно преобладающую, но раньше существовали одни только селахии и ганоидные; и в данном случае мы можем решить, повысились или деградировали рыбы в своей организации, только согласно с тем, какой критерий высоты организации мы пожелаем избрать. «
«
(Чарльз Дарвин «ПРОИСХОЖДЕНИЕ ВИДОВ ПУТЁМ ЕСТЕСТВЕННОГО ОТБОРА»)
волшебная палочка анафемы
Ты будешь прощён гаубицей
во фраке членораздельного целомудрия.
Ты посмеешь кошнотворной марионеточной кашей
заплести в свою бороду неприкосновенные запасы
и прославить своего недоношенного тушканчика.
Ты вылижешь копытце в привокзальном сортире.
А потом…….- когда на семи соборах
тебя предадут анафеме,-
ты станешь волшебной палочкой
с ядовитой волчьей иголкой на конце.
Ты — перелетишь….
……………………………
опрятный опыт суицида черноземья
В отборный чернозём
позём слезы и фортепьяно.
Но пьяный пахарь на подборке лески
опять сорвёт брусничные бурлески
и скинет на престол сапог кирзовый
в мокрой гальке.
Но пьяный пахарь украдёт у Фальконе
верёвку, мыло и одеколон……..
(И будет! Вавилон.)
усталости исторических блокад
Стойкие исламские солдатики
сломали протезы у фиолетовой рани.
Модильяни их журил апельсиновой стайкой,-
втыкал дротики в пролетающие товарники,
мыло стирал у реки,
плавил суставы в гробницу Тутанхамона.
Оборона нетленной плоти — сгущалась…
Оборона — становилась облачными карамельками.
мутное
Ближний рот и нахлебник Хлебникова…
Так блокада полирует сферу желудка,
так тепло из тёплого — плывёт в патефонные запонки
и… — не прощает.
Лишь — дымится. дымится. дымится.
рожать буду я, уважаемые литераторы
Литературные женишки,
книжки-побрякушки,
поэты-ракушки.
Купите мне простынку для родов;
скиньте с маятника лета.
Та (стратосферная и авторитарная) —
уже не изменит своих намерений стать лифтом,
шляющимся по моим гиенным венам.
барыга сказочный
(- Из морева в атласные ляжки барышень! -)
Так рассуждал барыга,
обижавший надувные трамваи.
Караван-сараи дрызгались кудрями светотеней
его изломанного характерного смеха.
Барыга скупал выживших в кислороде рыб
и выкруживал ими именины вождей…
Но больше всего он любил тугоплавкую ненависть.
морское солёное и земное сладкое
У рубикона силиконовых акул родня — менты.
И водоросли ждут стяженье жил и нервов. —
Им хочется маневров Н2О.
Окно у Посейдона — жаберный распах!
Окно у Посейдона — ветреный плавник!
Из них — никто не помнит,
как в домике (в Коломне) —
вкусны брюссельские кошачьи лапки
и негниющее мороженое спело……..
бесконец фильма
Конец фильмы!
Конец протуберанцам цапли,
что вы-ха-ха-лью лаяла на капли,
сочившиеся из ГУЛАГа…
Отвага и навага — не преодолимы!:
Когда кинотеатр зверью продыхает
в замочность скважин
сталь, отмычки
и крохотный бубенчик сердца.
террористическое женское против искусства!!!
Из бесконечного бюстгальтера упали —
улыбки, твёрдый знак на мягкий
и вертолёт, направленный на Третьяковку.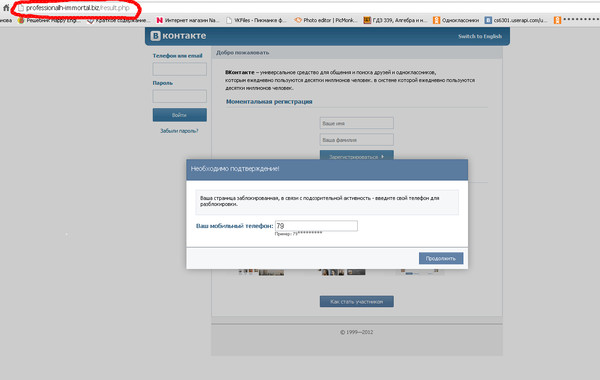
Сноровкой снайпера тогда распался атом
на купидонов, лелей и блядей,
намучивших юнцов янтарной гонореей.
А канарейки — что?
Пустяк и клюв в наморднике из трелей
… опять пристрелен, как облезлая дворняга…
… как Сукин Сын — Сергеич…… Александр.
про письма……
Крупнолистые калитки
свитки сабельных фаворитов
раздарили на письма.
Письма дрожали жалью погасшей пчелы:
маялись, татуировались,
кодировались от пьянства
невыносимой лёгкости медовухи.
электрическая зарисовка о сыне человеческом и некрофоне
Бочка на сносях Боттичелли
легла в качели — опрятная,
как свежепокрашенный покойник…
Как подоконник оступившийся
в свободную нишу суицида.
Коррида бычью жизни
утверждала математические поиски
Сына Человеческого! —
Но он не поддавался.-
Лишь двумя пальчиками исследовал розетку.
7 на 7
Семь дочерей на семи повешенных
зимуют,-
греются в горсточке оргазма.
Только “лёгкими фасонами” и пассажами
прерывают они своё зимовье,
чтобы постучать столом об кулак,
чтобы побрехать акафистом к Леониду Андрееву.
куций хвостик порося и волшебный волосок льва николаича
Куций звёздный хвостик порося,
прося варежки или электроэнергии,
превращался лицо-шарабан,
заискивая чан простуды и студня вселенной.
Но белое колено перемололо его
на бесчисленные волосики
из бороды-дуды потеплевшего Льва Николаича…
комплекс гердты и бестселлер: «зачемистый вопросник»
Зачемистый вопросник отвечал,
как на причал присела муза
с арбузами, листовками и песней
о краешке стола,
о попрошайной шали,
о необеденном бомже и перерыве.
Но это только отклоненье вектра;
но это Комплекс Гердты.
книжарь-мифодий о самодержавии канцелярских крыс
Книжарь, Мефодий!
Родин нет в печатности пера,
в початости рябин — нет горя. ..
..
Лишь частно-собственность
на полках процветает авторитаризмом,
которую утрируют ночами
мангусты — самодержавцы канцелярских крыс.
Как защитить страницу Вконтакте
Обновлено — 19 марта 2019 в 15:08
В связи с этим, каждый день хакеры взламывают сотни аккаунтов. Конечно, в большинстве случаев виноваты сами владельцы страниц (слабый пароль, пользование прокси-серверами, переходы на фишинговые сайты).
Злоумышленники легко подбирают пароли, создают фишинговые сайты и делают всё возможное, чтобы получить доступ к любым страницам. Ведь для них это выгодно: они продают аккаунты для коммерческих целей (в основном — раскрутка групп Вконтакте).
Поэтому защитить страницу Вконтакте от хакеров (взлома) необходимо каждому пользователю. И сейчас Вы узнаете как это сделать в 2017 году.
Как защитить страницу Вконтакте
Уверен, что у Вас он довольно простой, поэтому переходим в раздел «Настройки» и в первой вкладке указываем новый пароль:
Надёжный пароль должен состоять как из латинских символов, так и из цифр. Лучше всего, если несколько символов будут заглавными (например, хороший пароль — iniEstA109).
Лучше всего, если несколько символов будут заглавными (например, хороший пароль — iniEstA109).
2. Следующим действием будет настройка безопасности страницы. Из раздела «Настройки» переходим во вторую вкладку «Безопасность» и выполняем следующие действия:
Защита аккаунта в ВК
Таким образом, если кто-то сидит с Вашей страницы, то после этого действия его выкинет с аккаунта и ему придётся снова вводить логин и пароль. А так как Вы поменяли пароль, то больше он не войдёт на Вашу страницу.- Обязательно ставим галочку возле графы «Защита передаваемых данных»;
Тем самым злоумышленники не смогут перехватить Ваши данные через ненадёжные интернет-соединения (например, wi-fi в кафе).
Таким образом, для того, чтобы войти на Вашу страницу с другого браузера или устройства, необходимо будет ввести одноразовый код, который придёт на Ваш мобильный телефон.
После всех вышеописанных действий Ваша страница будет полностью защищена от взлома.

Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
PLOS ONE: Медицина и здравоохранение
Шуайци Го, Кристофер П. Гарнхэм, […], Питер Л. Дэвис
Казуя Кубояма, Акихиро Фудзикава, […], Масахару Нода
Масая Таяма, Масару Татено, […], Тошиказу Сайто
Ми Чжон Ли, Донг Хо Шин, […], Син-Ук Кан
Сунг-Хун Ли, Мухаммад Захур, […], Кан-Йелл Чой
Дженнифер Х. Стерн, Кёнми Ким, Джон Дж. Рэмси
Лука Пассамонти, Грэм Фэирчайлд, […], Эндрю Дж. Колдер
Изабелла Апаресида Салерно Пиментель, Каролина де Сикейра Палади, […], Клара Люсия Барбьери
Хуан Ши, Ифэй Чжан, […], Джи Хун
Хема Мартин-Мансо, Дхаммика Х. М. Л. П. Наваратна, […], Дэвид Д. Робертс
Бенедикте Менгель Перс, Сандип Кришна, […], Могенс Х. Йенсен
Сангита Дандекар, Цзянь Дин, […], Стэнли А. Кляйн
Программа| CyFy 2020 | ORF
Повестка дня — день 1
Понедельник, 12 октября 2020 г. Обратите внимание, что все часы указаны в индийском стандартном времени IST (GMT +5: 30)В политике, которой мы доверяем: разделение и цифровая взаимозависимость
Диалог Индия-АСЕАН, Трек 1.5 по кибервопросам (только по приглашению)
ВНУТРЕННЯЯ СЕССИЯ
Инаугурация и приветственное слово Sunjoy Joshi
Председатель, Observer Research Foundation
Самир Саран
Председатель CyFy, президент Observer Research Foundation
Триша Рэй
Сопредседатель CyFy, младший научный сотрудник, Observer Research Foundation
Адреса министров
Мулай Хафид Элалами
Достопочтенный министр промышленности, инвестиций, торговли и цифровой экономики Королевства Марокко
Тимо Харакка
Достопочтенный министр транспорта и коммуникаций Финляндии
Достопочтенный Джон Уиттингдейл OBE MP
Государственный министр по делам СМИ и данных
Инаугурационная речь
Рави Шанкар Прасад
Министр связи, электроники и информационных технологий, юстиции и юстиции
Возвращение суверена: определение границ в киберпространстве?
Когда-то подавляющее господство Соединенных Штатов над технологиями и потоками данных оспаривается, в частности, Китаем, но также и Европой и странами Азии с формирующимся рынком.Одновременно эти потоки стали неразрывно связаны с безопасностью и стабильностью государств и обществ. С призывами к новым инструментам, таким как D10, а также с растущими битвами по установлению стандартов и нормотворчеству новых технологий, наблюдаете ли вы возрождение Левиафана в киберпространстве?
Пропущенные подключения: конфликт из-за 5G
В 2020 году, когда мир остановился, боевые линии были обозначены по поводу тестирования и развертывания 5G.Поскольку все больше стран предпочитают дистанцироваться от Huawei и ZTE из-за своих уютных отношений с КПК, другие уже упираются в пятки, отказываясь запрещать иностранным поставщикам. На этой панели будут обсуждены стратегии, применяемые в разных географических регионах, чтобы сбалансировать императивы безопасности, устойчивого экономического роста и бинарных отношений между США и Китаем в отношении 5G.
Speedtalk о зрелости кибербезопасности, Баси фон Зольмс, директор Центра кибербезопасности Йоханнесбургского университета
Бэси фон Зольмс
Директор Центра кибербезопасности Йоханнесбургского университета
Реакции цепочки поставок: масштабные уязвимости?
В эпоху, отмеченную «деглобализацией» и «регионализацией», страны изучают не только торговые взаимозависимости, но и взаимозависимости в киберпространстве.От полной безопасности цепочки поставок компонентов до собственных стеков API и защиты данных — концепция безопасности в киберпространстве теперь охватывает каждую мелочь
Повестка дня — День 2
Вторник, 13 октября 2020 г. Обратите внимание, что все часы указаны в индийском стандартном времени IST (GMT +5: 30)Пандемия Прометея: инновации через кризис
Море фишинга: необходимость повышения киберустойчивости (по приглашению)
Основной доклад министров: «Технологии и пандемия» С.Исваран, почетный министр связи, информации и кибербезопасности, Сингапур
С. Исваран
Достопочтенный министр связи, информации и кибербезопасности, Сингапур
В поисках панацеи: технологии во времена кризиса
Беспрецедентный размах и влияние пандемии были встречены гонкой всех наций за разработку решений, от приложения для отслеживания контактов и хакатонов COVID до использования облачных вычислений и искусственного интеллекта для разработки вакцин.Что могут сделать правительства для создания благоприятной политической среды для безопасной разработки, тестирования, развертывания и использования этих новых решений?
У алтаря решения проблем? Цифровые права и интеграция после пандемии
Быстрое распространение технологий наблюдения и мониторинга после пандемии высветило вопросы, касающиеся конфиденциальности, доступа к цифровым технологиям и прав. Поскольку страны и компании принимают «решения», требующие от людей отказа от личной жизни для сохранения средств к существованию и мобильности, необходимо предостеречь от новых «ненормальных», которые мы можем создавать.Действительно ли права и безопасность — это дихотомия? Как мы можем развить нынешний кризис, чтобы создать режим «технология для всех» и «технология для всех»?
17:45 — 17:55
Speedtalk от Джейн Холл Лют
Президент и генеральный директор, SICPA North America
Планета Plandemic: борьба с Infodemic
От глав государств до террористов нет недостатка в субъектах, готовых манипулировать платформами для создания и распространения дезинформации и дезинформации.Пандемия Covid-19 переросла в существующую инфодемию, нанося дальнейший ущерб цифровой общественной сфере. Это глобальная проблема, и исследования показывают, что злоумышленники все чаще «навязывают» информационные операции друг друга. Хотя государства начали бороться с кризисом дезинформации и дезинформации, глобальные ответные меры остаются разрозненными и фрагментарными. Как государства, платформы и сообщества могут сбалансировать противоречия между национальными правовыми режимами и международными правовыми рамками в киберпространстве?
Диалог между Индией и США (только по приглашениям)
Повестка дня — день 3
Среда, 14 октября 2020 г. Обратите внимание, что все часы указаны в индийском стандартном времени IST (GMT +5: 30)iWork: равенство, занятость и идентичность в цифровой экономике
Future of Skills (только по приглашениям)
Основной доклад
Мариз Пейн
Достопочтенный министр иностранных дел Австралийского Союза
WeDigital: рабочие места и рабочая сила
На протяжении веков трудовые отношения строились вокруг общего рабочего пространства — реальность, в которой технологии, которым недавно помог коронавирус, быстро меняются.Каким будет будущее профсоюзов и переговоров с ростом числа цифровых рабочих и «облачных» офисов? Как в законодательстве следует признать новые формы коллективов и устранить информационную асимметрию между работниками и руководством?
Speedtalk
Рапеланг Рабана
Основатель и председатель Rekindle Learning
Общества нулевого дня: закодированный патриархат
За последние несколько лет в нескольких опросах был задокументирован интересный феномен: женщины склонны «доверять» ИИ меньше, чем мужчины, что, возможно, отражает интуитивное понимание того, что алгоритмы прогнозирования будут различать старые линии социального разлома, что убедительно продемонстрировали инструменты ИИ, которые выявили десятилетия дискриминации при приеме на работу.Достаточно ли внимания уделяется гендерным аспектам виртуального или полувиртуального рабочего пространства? Как укоренить понятие равенства в найме, рабочей среде и организационных процессах?
17:45 — 17:55
Speedtalk
Д-р Ахмед Эльмагармид
Исполнительный директор Катарского исследовательского института вычислительной техники
Расширяя границы индийской технологии: женщины на передовой
Технологии — неотъемлемая часть социально-экономического роста Индии.В настоящее время Индия является свидетелем двух поворотных феноменов, ориентированных на женщин: во-первых, наблюдается беспрецедентное включение женщин в индийскую экономику благодаря технологическим платформам, таким как India Stack. Во-вторых, женщины все больше расширяют границы развития технологий. Каким образом правительство, предприниматели, новаторы и политики могут объединиться, чтобы укрепить эту тенденцию и добиться большего роста, основанного на инновациях коренных народов?
Модератор: Ниша Холла
Научный сотрудник Центра клеточных и молекулярных платформ
Киран Мазумдар Шоу
Председатель, Biocon Ltd.
Ниврути Рай
Глава страны, Intel Индия
Повестка дня — день 4
Четверг, 15 октября 2020 г. Обратите внимание, что все часы указаны в индийском стандартном времени IST (GMT +5: 30)Байт демократии: от народа, от народа, против народа?
Основной доклад
Thomas Heilmann
Член парламента, член комитета по цифровой повестке дня, Бундестаг
Сессия завтрака: Индо-тихоокеанское сотрудничество в области критических технологий
Speedtalk: «Дипфейки и демократия»
Ашиш Джайман
Директор по технологическим операциям, Microsoft
Избирательная урна и чудовище: безопасность наших выборов
Открытые общества становятся все более уязвимыми для злоумышленников.Информационные потоки и общественное мнение; политические кампании и отдельные лица; системы регистрации и голосования; и соответствующая избирательная инфраструктура находятся под угрозой из-за скоординированных операций влияния, как внутренних, так и зарубежных. Какие правовые рамки, технологические инструменты и гарантии могут принять демократии для защиты неприкосновенности кабины для голосования?
Основной доклад министров
Томаш Петршичек
Министр иностранных дел Чешской Республики
Бегущий вслепую: перегрузка технологий
В определенных условиях люди по своей природе доверяют решениям, принимаемым машинами, а не решениям, принимаемым людьми, иногда за счет независимого человеческого суждения.Не пора ли оценить, где технологии стали препятствием, а не решением? Как мы можем повысить устойчивость к злонамеренному вмешательству и вредоносным нарративам на индивидуальном и организационном уровне?
Беседа: будущее цифровой Европы
Повестка дня — день 5
Пятница, 16 октября 2020 г. Обратите внимание, что все часы указаны в индийском стандартном времени IST (GMT +5: 30)Techreation: создание, потребление и раскрытие ценности посредством экономики данных
Speedtalk Вик Кроун, генеральный директор Callaghan Innovation
Вик Крон
Генеральный директор Callaghan Innovation
Сотворение во времена COVID: повествование в темные времена
Для миллионов из нас в этом году наши экраны стали нашим окном в мир, и создатели контента — от рекламодателей до режиссеров и писателей — столкнулись с проблемой продолжения рассказа захватывающих историй, даже когда все было выключено.Как искусство и содержание повествования адаптировались к этим странным временам?
Модератор: Митали Мукерджи
Сотрудник, Исследовательский фонд наблюдателей
Каран Аншуман
Режиссер и продюсер
Хари Кришнан
Основатель, Culture Drum
Доверие к транзакциям: устранение узких мест при внедрении цифровых платежей
По мере того, как страны закрываются, цифровой рост, особенно в странах с развивающейся экономикой, стремительно растет, поскольку города обращаются к онлайн-услугам, от еды и работы до развлечений и досуга.Цифровые платежи будут играть все более важную роль в формирующихся экономиках, ориентированных прежде всего на цифровые технологии. Однако в некоторых случаях появление альтернативных платежных систем привело к отсутствию доверия среди потребителей, а также к проблемам для регулирующих органов в незнакомых водах. Как минимизировать трения между регуляторами, потребителями и игроками в сфере финансовых технологий? Какие появляются новые модели регулируемых инноваций в пространстве цифровых платежей?
Смена каналов: развлечения после пандемии
В разгар пандемии, даже когда кинозалы по всему миру опустели, количество подписчиков на сервисы потокового видео росло, продажи видеоигр росли, а музыкальные исполнители проводили яркие интерактивные онлайн-концерты для аудитории, которая намного превосходила по численности зрителей, живущих в реальных залах. .Наблюдаем ли мы медленную смерть традиционных СМИ или новую эру создания кросс-медиа контента?
Модератор: Виван Шаран
Партнер, Koan Advisory Group
Вани Трипати Тико
Член правления, Центральный совет сертификации фильмов
Шекхар Капур
Режиссер
Основной доклад
Аджит Пай
Председатель, Федеральная комиссия по связи, США
Публичные сферы, частные правила: открытый дискурс в киберпространстве
За прошедшие годы онлайн-платформы стали ключевыми проводниками важных публичных дискуссий.Следовательно, общественные сферы находятся под влиянием частных корпораций, поскольку дискурсы на таких платформах подчиняются нормативным рамкам, администрируемым их провайдерами. Возникает вопрос, имеют ли корпорации право модерировать публичные дискурсы, в том числе во внешних юрисдикциях, путем применения своих собственных субъективных параметров? Какие меры следует предпринять, чтобы эти обсуждения оставались по-настоящему свободными и открытыми? Социальные сети вышли из-под контроля?
Модератор: Миши Чаудхари
Основатель и юридический директор, SFLC
Дипаян Гош
Основатель лаборатории локализации
Виктория Иванченко
Главный редактор, PICREADI
Шита Лакшми
Исполнительный директор, Фонд Тифа
Speedtalk
Аста Капур
Соучредитель Aapti
История двух городов: конкурирующие взгляды на управление данными
Крупнейшие технологические гиганты нашего времени не продают физический «продукт», а привлекают внимание своих пользователей.В разных географических регионах применяются разные подходы к балансированию конкурирующих требований государства, гражданина / пользователя и промышленности. Некоторые из них привели к враждебным отношениям между этими тремя, в то время как другие стремятся к синергии. На этой панели будут рассмотрены передовые практики, вытекающие из этих конкурирующих моделей управления данными.
Заключительное заседание
Самир Саран
Председатель CyFy, президент Observer Research Foundation
Триша Рэй
Сопредседатель CyFy, младший научный сотрудник, Observer Research Foundation
Нормативно-правовая база для цифровой экономики на триллион долларов (только по приглашению)
Диалог между ЕС и Индией (только по приглашениям)
Таймс-сквер, серия
12–16 октября 2020 г. — Обратите внимание, что все часы указаны по индийскому стандартному времени IST (GMT +5: 30)
Джеймс Льюис
Старший вице-президент, CSIS
Меган Стифель
Старший научный сотрудник, Cyber Statecraft Initiative
Дипаян Гош
Соруководитель проекта «Цифровые платформы и демократия» Гарвардской школы Кеннеди
Миши Чоудхари
Основатель и юридический директор, SFL
Мегха Арора
Инженер по гражданским свободам, Palantir Technologies
% PDF-1.4 % 4642 0 объект > эндобдж xref 4642 236 0000000016 00000 н. 0000014047 00000 п. 0000014328 00000 п. 0000014374 00000 п. 0000014412 00000 п. 0000015389 00000 п. 0000015428 00000 п. 0000016225 00000 п. 0000017124 00000 п. 0000017969 00000 п. 0000018773 00000 п. 0000019625 00000 п. 0000020443 00000 п. 0000020842 00000 п. 0000021306 00000 п. 0000022096 00000 п. 0000023099 00000 н. 0000025750 00000 п. 0000025815 00000 п. 0000025888 00000 п. 0000025948 00000 п. 0000026031 00000 п. 0000028205 00000 п. 0000028540 00000 п. 0000028626 00000 п. 0000031848 00000 п. 0000032218 00000 п. 0000032522 00000 п. 0000032722 00000 п. 0000032830 00000 н. 0000032967 00000 п. 0000033167 00000 п. 0000033276 00000 п. 0000033413 00000 п. 0000033716 00000 п. 0000033824 00000 п. 0000033961 00000 п. 0000034348 00000 п. 0000034439 00000 п. 0000034576 00000 п. 0000034963 00000 п. 0000035054 00000 п. 0000035191 00000 п. 0000035673 00000 п. 0000035724 00000 п. 0000044809 00000 п. 0000044998 00000 н. 0000045048 00000 п. 0000045438 00000 п. 0000045921 00000 п. 0000045972 00000 п. 0000048174 00000 п. 0000049039 00000 н. 0000049547 00000 п. 0000049598 00000 п. 0000050198 00000 п. 0000050708 00000 п. 0000050759 00000 п. 0000051340 00000 п. 0000051547 00000 п. 0000051597 00000 п. 0000051925 00000 п. 0000052149 00000 п. 0000052199 00000 п. 0000052530 00000 н. 0000053445 00000 п. 0000054308 00000 п. 0000054810 00000 п. 0000054861 00000 п. 0000062062 00000 п. 0000065573 00000 п. 0000084745 00000 п. 0000110395 00000 н. 0000130368 00000 н. 0000160492 00000 п. 0000176712 00000 н. 0000183186 00000 н. 00001
00000 н. 0000192447 00000 н. 0000193211 00000 н. 0000199153 00000 н. 0000205851 00000 н. 0000211575 00000 н. 0000221617 00000 н. 0000227958 00000 н. 0000244956 00000 н. 0000264659 00000 н. 0000286001 00000 п. 0000286077 00000 н. 0000286179 00000 н. 0000286298 00000 н. 0000286422 00000 н. 0000286580 00000 н. 0000286816 00000 н. 0000286967 00000 н. 0000287079 00000 п. 0000287215 00000 н. 0000287392 00000 н. 0000287510 00000 н. 0000287622 00000 н. 0000287812 00000 н. 0000287930 00000 н. 0000288042 00000 н. 0000288220 00000 н. 0000288338 00000 п. 0000288450 00000 н. 0000288638 00000 п. 0000288756 00000 н. 0000288868 00000 н. 0000289071 00000 н. 0000289283 00000 п. 0000289414 00000 н. 0000289526 00000 н. 0000289753 00000 н. 0000289894 00000 н. 00002
00000 н. 00002 00000 н. 00002 00000 н. 0000200000 н. 00002 00000 н. 00002 00000 н. 00002
00000 н. 0000200000 н. 00002
Реакция культур на засуху и интерпретация адаптации
Ангус Дж. Ф. и Монкур М. В. (1977) Водный стресс и фенология пшеницы. Aust J Agric Res 28: 177–181
Google Scholar
Бартельс Д. и Нельсон Д. (1994) Подходы к повышению устойчивости к стрессу с использованием молекулярной генетики, клетки растений и среда 17: 659–667
Google Scholar
Basnayake J, Copper M, Ludlow MM and Henzell RG (1994) Объединение вариаций осмотической способности регулировок между выбранным диапазоном зернового сорго ( Sorghum bicolor L.Moench) линий. Полевые культуры Res 38: 147–155
Статья Google Scholar
Бидингер FR, Махалакшми В. и Рао GDP (1987) Оценка засухоустойчивости жемчужного проса ( Pennisetum americanum (L.) Leeke). I. Факторы, влияющие на урожайность в условиях стресса. Aust J Agric Res 38: 37–48
Google Scholar
Белески Р.Л. (1993) Гидролиз фруктана вызывает расширение лепестков эфемерного цветка лилейника.Физиология растений 103: 213–219
PubMed Google Scholar
Блюм А. (1988) Селекция растений в стрессовых условиях. CRC Press, Бока-Ратон, Флорида, 208 стр.
Google Scholar
Блюм А. (1992) Выбор для устойчивого производства в условиях дефицита воды. Internat Crop Sci I, Crop Sci Soc Am, Мэдисон, стр. 343–347
Google Scholar
Blum A и Ebercon A (1981) Стабильность клеточной мембраны как мера устойчивости пшеницы к засухе и жаре. Crop Sci 21: 43–47
Google Scholar
Блюм А. и Аркин Г.Ф. (1984) Рост корней сорго и водопотребление в зависимости от водоснабжения и продолжительности роста. Полевые культуры Res 9: 131–142
Статья Google Scholar
Блюм А. и Ричи Дж. Т. (1984) Влияние содержания воды на поверхности почвы на распределение корней сорго в почве.Полевые культуры Res 8: 169–176
Article Google Scholar
Блюм А. и Джонсон Дж. В. (1992) Перенос воды из корней в сухую почву и влияние на водные отношения и рост пшеницы. Растения и почва 145: 141–146
Google Scholar
Блюм А. и Пнуэль Ю. (1990) Физиологические признаки, связанные с засухоустойчивостью сортов пшеницы в средиземноморской среде.Aust Jour Agric Res 41: 799–810
Google Scholar
Блюм А., Аркин Г.Ф. и Джордан В.Р. (1977) Морфогенез и рост корней сорго. I. Влияние генов зрелости. Crop Sci 17: 149–153
Google Scholar
Блюм А., Голан Г., Майер Дж., Синмена Б. и Обилана Т. (1992) Сравнительная продуктивность и реакция на засуху полутропических гибридов и открытоопыляемых сортов сорго.J Agric Sci Camb 118: 29–36
Google Scholar
Блюм А., Джонсон Дж. В., Рамсер Е. Л. и Толлнер Е. В. (1991) Влияние высыхания верхнего слоя почвы и возможного негидравлического корневого сигнала на рост и урожайность пшеницы. J Exp Bot 42: 1225–1231
Google Scholar
Блюм А., Майер Дж. И Голан Г. (1988) Влияние количества зерен (размера поглотителя) на активность источников и их водные отношения в пшенице.J Exp Bot 39: 106–114
Google Scholar
Блюм А., Майер Дж. И Гозлан Г. (1983) Связь между производством растений и некоторыми физиологическими компонентами засухоустойчивости пшеницы. Растительные клетки и окружающая среда 6: 219–225
Google Scholar
Блюм А., Рамайя С., Канемасу Е.Т. и Полсен Г.М. (1990) Восстановление пшеницы после стресса засухи на стадии развития кущения.Полевые культуры Res 24: 67–85
Article Google Scholar
Блюм А., Шпилер Л., Голан Г. и Майер Дж. (1990b) Стабильность урожайности и температура растительного покрова генотипов пшеницы при стрессе засухи. Полевые культуры Res 22: 289–296
Статья Google Scholar
Блюм А., Синмена Б. и Зив О. (1980) Оценка скрининговых тестов на засухоустойчивость семян и проростков пшеницы.Euphytica 29: 727–736
Google Scholar
Блюм А., Синмена Б., Майер Дж., Голан Г. и Шпилер Л. (1994) Мобилизация запаса стебля поддерживает наполнение зерна пшеницы при тепловом стрессе. Aust J Plant Physiol 21: 771–781
Google Scholar
Боланос Дж. И Эдмидс Г.О. (1991) Значение отбора по осмотическому потенциалу тропической кукурузы. Agron J 83: 948–956
Google Scholar
Boyle MG, Boyer JS и Morgan PW (1991) Вливание стеблей жидкой питательной среды предотвращает репродуктивную недостаточность кукурузы при низком водном потенциале. Crop Sci 31: 1246–1252
Google Scholar
Бирн П.Ф., Боланос Дж., Эдмиадс Г.О. и Итон Д.Л. (1995) Выгоды от отбора в условиях засухи по сравнению с тестированием на нескольких участках в родственных популяциях тропической кукурузы. Crop Sci 35: 63–69
Google Scholar
Casal JJ (1988) Влияние качества света на внешний вид побегов разного порядка у пшеницы ( Triticum aestivum ). Ann Appl Biol 112: 167–173
Google Scholar
Ceccarelli S и Grando S (1991) Селекционная среда и экологическая чувствительность ячменя. Euphytica 57: 157–167
Google Scholar
Chandler PM и Robertson M (1994) Экспрессия генов, регулируемая абсцизовой кислотой, и ее связь с устойчивостью к стрессу.Ann Rev Plant Physiol и Mol Biol 45: 113–141
Статья Google Scholar
Чавес М.М. (1991) Влияние дефицита воды на ассимиляцию углерода — обзорная статья. J Exp Bot 42: 1–16
Google Scholar
Craufurd PQ, Flower DJ и Peacock JM (1993) Влияние стресса от жары и засухи на сорго ( Sorghum bicolor ). 1. Развитие метелки и внешний вид листьев.Exp Agric 29: 61–76
Google Scholar
Дэвис В.Дж., Тардье Ф. и Трехо С.Л. (1994) Как химические сигналы работают в растениях, которые растут в высыхающей почве. Plant Physiol 104: 309–314
PubMed Google Scholar
Донателли М., Хаммер Г.Л. и Вандерлип Р.Л. (1992) Влияние генотипа и ограничения воды на фенологию, рост и эффективность транспирации у зернового сорго Crop Sci 32: 781–786
Google Scholar
Dwyer LM и Stewart DW (1987) Влияние фотопериода и водного стресса на рост, урожайность и скорость развития ячменя, измеряемые в тепловых единицах. Can J Plant Sci 67: 21–34
Google Scholar
Элалауи А.К., Симмонс С.Р. и Крукстон Р.К. (1992) Распределение фотоассимилята основными побегами и невыжившими побегами ячменя Crop Sci 32: 1233–1237
Google Scholar
Фаркуар Г. Д., Вонг С. К., Эванс Дж. Р. и Хубик К. Т. (1989) Фотосинтез и газообмен. В: Джонс Х.Г., Флауэрс Т.Дж. и Джонс М.Б. (ред.) «Растения под стрессом», стр. 47–69. Издательство Кембриджского университета, Кембридж
Google Scholar
Flower DJ, Rani AU и Peacock JM (1990) Влияние осмотической регулировки на рост, устьичную проводимость и перехват света контрастных линий сорго в суровых условиях. Aust J Plant Physiol 17: 91–105
Google Scholar
Гэррити Д.П., Салливан С.Ю. и Уоттс Д.Г. (1984) Изменения устьичной и фотосинтетической реакции зерна сорго на стресс от влаги на разных стадиях роста. Crop Sci 24: 441–446
Google Scholar
Garrity DP, Vidal ET и O’Toole JC (1986) Управление сопротивлением транспирации метелки для увеличения фертильности колосков во время водного стресса на стадии цветения. Crop Sci 26: 789–795
Google Scholar
Grantz DA (1990) Реакция растений на атмосферную влажность. Растительная клетка и окружающая среда 13: 667–679
Google Scholar
Гупта А.С. и Берковиц Г.А. (1987) Осмотическая регулировка, объем симпласта и нестоматически опосредованное подавление водного стресса фотосинтеза у пшеницы. Физиология растений 85: 1040–1047
Google Scholar
Холл А.Е., Ричардс Р.А., Кондон А.Г., Райт Г.К. и Фаркуар Г.Д. (1994) Дискриминация изотопов углерода и селекция растений Породы растений.Ред. 12: 81–113
Google Scholar
Hamblin J (1993) Концепция идеотипа: полезная или устаревшая? Междунар. Crop Sci I, Crop Sci Soc Am, Madison, стр. 589–597
Google Scholar
Havaux M (1992) Стрессоустойчивость фотосистемы-II in vivo — антагонистические эффекты воды, тепла и стрессов фотоингибирования Plant Physiol 100: 424–432
Google Scholar
Hendrix JH, Linden JC, Smith DH, Ross CW и Park IK (1986) Связь метаболизма фруктана перед цветением с количеством зерен у озимой пшеницы ( Triticum aestivum L.). Aust J Plant Physiol 13: 391–398
Google Scholar
Хенсон И.Е. и Махалакшми В. (1985) Доказательства контроля метелками устьичного поведения у испытывающих водный стресс растений жемчужного проса. Полевые культуры Res 11: 281–290
Article Google Scholar
Johnson DA и Asay KH (1993) Точка зрения — выбор для улучшения реакции на засуху трав в холодный сезон. J Range Manage 46: 194–202
Google Scholar
Кан Т.Л., Фендер С.Р., Брей Э.А. и Оконнелл М.А. (1993) Характеристика экспрессии генов томатов, регулируемых засухой и абсцизовой кислотой, у засухоустойчивых видов Lycopersicon-pennellii . Физиология растений 103: 597–605
PubMed Google Scholar
King RW и Evans LT (1977) Подавление цветения у Lolium temulentum водным стрессом: роль абсцизовой кислоты. Aust J Plant Physiol 4: 225–233
Google Scholar
Кошкин Е.И., Тарарина В.В. (1990) Различия в соотношении источник-поглотитель в пшенице и их связь с урожаем зерна и содержанием абсцизовой кислоты. Plant Physiol Biochem 28: 609–616
Google Scholar
Kriedemann PE (1986) Стоматологические и фотосинтетические ограничения роста листьев. Aust J Plant Physiol 13: 15–31
Google Scholar
Леунинг Р., Кондон А.Г., Дунин Ф.Х., Зегелин С. и Денмид О.Т. (1994) Перехват дождя и испарение из почвы под пологом пшеницы. Agr Forest Meteorol 67: 221–238
Статья Google Scholar
Lopezcastaneda C, Richards RA и Farquhar GD (1995) Различия в ранней силе роста между пшеницей и ячменем.Crop Sci 35: 472–479
Google Scholar
Lu ZM, Radin JW, Turcotte EL, Percy R и Zeiger E (1994) Высокая урожайность передовых линий хлопка пима связана с более высокой устьичной проводимостью, уменьшенной площадью листьев и более низкой температурой листьев Physiol. Завод 92: 266–272
Артикул Google Scholar
Mahalakshmi V, Alargarswamy G и Bidinger FR (1983) Связь между цветением и сниженной чувствительностью устьиц к водному стрессу у жемчужного проса (Pennisetum americanum (L.) Лик. Энн Бот 52: 641–648
Google Scholar
Малик Р.С., Дханкар Дж. С. и Тернер Н. С. (1979) Влияние дефицита влаги в почве на рост корней сеянцев хлопчатника. Растения и почва 53: 109–112
Google Scholar
Мартин Дж. Х. (1930) Сравнительная засухоустойчивость сорго и кукурузы. Agron J 22: 993–1003
Google Scholar
Макгоуэн М., Бланч П., Грегори П. Дж. И Хейкок Д. (1984) Водные отношения озимой пшеницы. 5. Корневая система и осмотическая регулировка в зависимости от испарения урожая. J Agric Sci 102: 415–425
Google Scholar
Мейер Дж., Скмитт Дж. М. и Бонерт Х. Дж. (1990) Прямой скрининг небольшого генома: оценка величины изменений экспрессии генов растений во время адаптации к высокому содержанию соли. Мол и Дженерал Генет 224: 347–356
Google Scholar
Mian MAR и Nafziger ED (1994) Влияние размера семян и водного потенциала на прорастание и рост проростков озимой пшеницы. Crop Sci 34: 169–171
Google Scholar
Mita SK и Suzukifujii NK (1995) Сахар-индуцируемая экспрессия гена бета-амилазы в Arabidopsis thaliana Plant Physiol 107: 895–904
PubMed Google Scholar
Morgan JM (1977) Изменения диффузионной проводимости и водного потенциала растений пшеницы до и после цветения. Aust J Plant Physiol 4: 75–86
Google Scholar
Morgan JM (1983) Осморегуляция как критерий отбора засухоустойчивости пшеницы. Aust J Agric Res 34: 607–613
Google Scholar
Morgan JM (1992) Осмотические компоненты и свойства, связанные с генотипическими различиями в осморегуляции у пшеницы.Aust J Plant Physiol 19: 67–76
Google Scholar
Morgan JM (1995) Рост и урожай линий пшеницы с различной осморегулирующей способностью при высоком дефиците воды в почве в сезоны с изменяющейся потребностью в испарении. Полевые культуры Res 40: 143–152
Статья Google Scholar
Morgan JM и Condon AG (1986) Использование воды, урожай зерна и осморегуляция пшеницы.Aust J Plant Physiol 13: 523–532
Google Scholar
Морган Дж. М., Родригесмарибона Б. и Найтс Э. Дж. (1991) Адаптация к дефициту воды в селекционных линиях нута путем осморегуляции — взаимосвязь с урожайностью зерна на поле. Полевые культуры Res 27: 61–70
Google Scholar
Нуден Л.Д. и Летэм Д.С. (1993) Метаболизм цитокининов и передача сигналов в растении сои.Aust J Plant Physiol 20: 639–653
Google Scholar
Oosterhuis DM и Cartwright PM (1983) Дифференциация колосов и выживаемость цветков у яровой пшеницы под воздействием водного стресса и фотопериода. Crop Sci 23: 711–717
Google Scholar
Пикок Дж. М., Миллер В. Б., Мацуда К. и Робинсон Д. Л. (1990) Роль теплового опоясания в ранней гибели проростков сорго.Crop Sci 30: 138–143
Google Scholar
Пинтер П.Дж., Зиполи Дж., Регинато Р.Дж., Джексон Р.Д., Идсо С.Б. и Хохман Дж.П. (1990) Температура растительного покрова как индикатор разницы в использовании воды и урожайности среди сортов пшеницы. Управление водных ресурсов сельского хозяйства 18: 35–48
Google Scholar
Quarrie SA (1993) Понимание реакции растений на стресс и селекция улучшили стрессоустойчивость — разрыв между поколениями.В: Close TJ and Bray EA (eds) Ответы растений на клеточное обезвоживание во время стресса окружающей среды. Актуальные темы физиологии растений. Am Soc of Plant Physiol Series 10: 224–245
Рид Дж. Дж., Эсей К. Х. и Джонсон Д. А. (1993) Дивергентный отбор для дискриминации изотопов углерода в пыреях хохлатых. Can J Plant Sci 73: 1027–1035
Google Scholar
Риз Д., Сэйр К., Асеведо Э., Нава Санчес Т., Лу З., Зейгер Э. и Лимон А. (1993) Температура растительного покрова пшеницы: взаимосвязь с урожайностью и потенциалом как метод отбора ранних поколений.Wheat Special Rep 10, СИММИТ, Мексика
Google Scholar
Рейнольдс М.П., Балота М., Дельгадо М.И.Б., Амани И. и Фишер Р.А. (1994) Физиологические и морфологические признаки, связанные с урожаем яровой пшеницы в условиях горячего орошения. Aust. J. Plant Physiol 21: 717–730
Google Scholar
Ричардс Р.А. (1992) Повышение засоления зерновых культур — стоит ли это.Растения и почва 146: 89–98
Google Scholar
Родригесмарибона Б., Тенорио Дж. Л., Конде Дж. Р. и Айерб Л. (1992) Корреляция между урожайностью и осмотической регулировкой гороха ( Pisum sativum L.) при стрессе засухи. Полевые культуры Res 29: 15–22
Google Scholar
Сантамария Дж. М., Ладлоу М. М. и Фукаи С. (1990) Вклад осмотической корректировки в урожай зерна у сорта Sorghum-bicolor (L) Moench в условиях ограниченного количества воды.1. Водный стресс перед цветением. Aust J Agric Res 41: 51–65
Google Scholar
Schnyder H (1993) Роль хранения и перераспределения углеводов в отношениях источник-поглотитель пшеницы и ячменя во время налива зерна — обзор. Новый Фитол 123: 233–245
Google Scholar
Schussler JR и Westgate ME (1994) Увеличение запасов ассимилятов не предотвращает аборта ядра кукурузы при низком водном потенциале.Crop Sci 34: 1569–1576
Google Scholar
Sedgley RH (1991) Оценка идеотипа Дональда через 21 год. Полевые культуры Res 26: 93–112
Google Scholar
Seemann JR и Sharkey TD (1987) Влияние абсцизовой кислоты и других ингибиторов на фотосинтетическую способность и биохимию ассимиляции CO2. Физиология растений 84: 696–700
Google Scholar
Зееманн Дж. Р., Даунтон В. Дж. С. и Берри Дж. А. (1986) Температура и осмотический потенциал листьев как факторы адаптации фотосинтеза к высокой температуре. в пустынных растениях. Физиология растений 80: 926–930
Google Scholar
Sharp RE (1990) Сравнительная чувствительность роста корней и побегов и физиология к низким водным потенциалам. Важность корня для коммуникации в ответ на стресс окружающей среды. Регламент роста растений Британского общества, Оксфорд, понедельник, 21: 29–44
Google Scholar
Шимши Д. и Эфрат Дж. (1975) Устьевое поведение сортов пшеницы в отношении их транспирации, фотосинтеза и урожайности. Agron J 67: 326–329
Google Scholar
Синклер Т.Р. и Ладлоу М.М. (1986) Влияние водоснабжения почвы на водный баланс четырех тропических зернобобовых культур. Aust J Plant Physiol 13: 329–341
Google Scholar
Smucker AJM и Aiken RM (1992) Динамические реакции корней на дефицит воды.Почвоведение 154: 281–289
Google Scholar
Снапп С.С. и Шеннан С. (1992) Влияние засоления на рост корней и динамику гибели томатов, Lycopersicon esculentum Mill. Новый Фитол 121: 71–79
Google Scholar
Стюарт Г.Р. (1989) Иссушение, ангидробиоз и выживаемость. 1989. В: Джонс Х.Г., Флауэрс Т.Дж. и Джонс М.Б. (ред.) Растения под стрессом, стр. 115–130.Издательство Кембриджского университета, Кембридж
Google Scholar
Стокман Ю.М., Фишер Р.А. и Бриттен Э.Г. (1983) Обеспечение ассимиляцией и развитие цветков в колосе пшеницы ( Triticum aestivum L.). Aust J Plant Physiol 10: 585–594
Google Scholar
Тангпремсри Т., Фукаи С., Фишер К.С. и Хенцелл Р.Г. (1991) Генотипические вариации в осмотическом регулировании в зерне сорго.2. Связь с некоторыми признаками роста. Aust J Agric Res 42: 759–767
Google Scholar
Tetteroo FAA, Peters AHL, Hoekstra FA, Vanderplas LHW и Hagendoorn MJM (1995) ABA снижает дыхание и метаболизм сахара в развивающихся эмбриоидах моркови ( Daucus carota L.). J Plant Physiol 145: 477–482
Google Scholar
Trewavas AJ и Jones HG (1991) Оценка роли ABA в развитии растений.В: Дэвис У. Дж. И Джонс Х. Г. (ред.) Физиология и биохимия абсцизовой кислоты, стр. 169–188. Bios Sci Publ Oxford UK
Google Scholar
Тернер NC, O’Toole JC, Cruz RT, Yambao EB, Ahmad S, Namuco OS и Dingkhun M (1986) Реакция семи различных сортов риса на дефицит воды. II. Осмотическая регулировка, эластичность листа, удлинение листа, гибель листа, устьичная проводимость и фотосинтез. Полевые культуры Res 13: 273–286
Article Google Scholar
Waters SP, Martin P и Lee BT (1984) Влияние сахарозы и абсцизовой кислоты на определение числа зерен пшеницы. J Exp Bot 35: 829–840
Google Scholar
Westgate ME и Boyer JS (1985) Осмотическая регулировка и ингибирование роста корня листьев и шелка при низком потенциале воды у кукурузы. Planta 164: 540–549
Google Scholar
White RH, Engelke MC, Morton SJ и Ruemmele BA (1992) Конкурентное поддержание тургора овсяницы высокорослой. Crop Sci 32: 251–256
Google Scholar
Wright GC и Smith RCG (1983) Различия между двумя генотипами сорго в адаптации к стрессу засухи. II. Поглощение воды корнями и водопотребление. Aust J Agric Res 34: 627–636
Google Scholar
Zinselmeier C, Westgate ME и Jones RJ (1995) Зерно, установленное на низкий водный потенциал, не зависит от соотношения источник / поглотитель в кукурузе.Crop Sci 35: 158–163
Google Scholar
1 | Ли Р.К., Фейнбаум Р.Л. и Амброс V: К. elegans гетерохронный ген lin-4 кодирует малые РНК с антисмысловыми комплементарность лин-14. Клетка. 75: 843–854. 1993. Просмотр статьи: Google Scholar: PubMed / NCBI | |
2 | Рейнхарт Б.Дж., Слэк Ф.Дж., Бассон М, Паскинелли А.Е., Bettinger JC, Rougvie AE, Horvitz HR и Рувкун Г. 21-нуклеотидная РНК let-7 регулирует время развития у Caenorhabditis elegans.Природа. 403: 901–906. 2000. Просмотр статьи: Google Scholar: PubMed / NCBI | |
3 | Ремсбург К., Конрад К., Сампило Н.Ф. и Сонг JL: Анализ функций микроРНК. Методы Cell Biol. 151: 323–334. 2019. Просмотр статьи: Google Scholar: PubMed / NCBI | |
4 | Suzuki H, Gabrielson E, Chen W, Анбажаган Р., ван Инглэнд М., Герман Дж. Г., Бейлин С.Б. и Вейенберг М.П .: Геномный скрининг генов, активируемых деметилированием и гистоном ингибирование деацетилазы при колоректальном раке человека.Нат Жене. 31: 141–149. 2002. Просмотр Статья: Google Scholar: PubMed / NCBI | |
5 | Вишной А. и Рани С. Биогенез MiRNA и регулирование болезней: обзор. Методы Мол биол. 1509: 1–10. 2017. Просмотр статьи: Google Scholar: PubMed / NCBI | |
6 | Линтнер Н.Г. и Кейт Дж.Х.Д .: Регулирование рибосома: в центре внимания темная материя РНК. Mol Cell. 54: 1-2. 2014 г. Просмотр статьи: Google Scholar: PubMed / NCBI | |
7 | Браннан С.И., Дис Э.С., Инграм Р.С. и Тилгман С.М.: Продукт гена h29 может функционировать как РНК.Mol Cell Biol. 10: 28–36. 1990. Просмотр статьи: Google Scholar: PubMed / NCBI | |
8 | Спиццо Р., Алмейда М.И., Коломбатти А и Калин GA: длинные некодирующие РНК и рак: новые рубежи трансляционные исследования? Онкоген. 31: 4577–4587. 2012. Просмотр статьи: Google Scholar: PubMed / NCBI | |
9 | Арриага-Канон C, Фонсека-Гусман Y, Вальдес-Кесада С., Арсате-Мехиа Р., Герреро Дж. И Ресиллас-Тарга Ф: Длинная некодирующая РНК способствует полной активации взрослого гена. экспрессия в курином α-глобиновом домене.Эпигенетика. 9: 173–181. 2014. Просмотреть статью: Google Scholar: PubMed / NCBI | |
10 | Бергманн Дж. Х. и Спектор Д. Л.: Лонг некодирующие РНК: Модуляторы ядерной структуры и функции. Curr Opin Cell Boil. 26: 10–18. 2014. Просмотр статьи: Google Scholar | |
11 | Майорос У.Х. и Олер У.: Пространственный предпочтения микроРНК-мишеней в 3′-нетранслируемых областях. BMC Геномика. 8: 1522007.Просмотр статьи: Google Scholar: PubMed / NCBI | |
12 | Кабили М.Н., Трапнелл С., Гофф Л., Козиол М., Tazon-Vega B, Regev A и Rinn JL: Интегративная аннотация человеческого большие межгенные некодирующие РНК обнаруживают глобальные свойства и конкретные подклассы. Genes Dev. 25: 1915–1927. 2011. Просмотр статьи: Google Scholar: PubMed / NCBI | |
13 | Du Z, Fei T, Verhaak RG, Su Z, Zhang Y, Brown M, Chen Y и Liu XS: интегративный геномный анализ показывает клинически значимые длинные некодирующие РНК при раке человека.Nat Struct Mol Biol. 20: 908–913. 2013. Просмотр статьи: Google Scholar: PubMed / NCBI | |
14 | Чжоу Цюй, Ю Цюй, Гонг Y, Лю Цзинь, Сюй Х, Ван И и Shi Y: Построение сети днРНК-миРНК-мРНК для определения регуляторные роли днРНК при псориазе. Exp Ther Med. 18: 4011–4021. 2019.PubMed / NCBI | |
15 | Донг З, Чжан А, Лю С., Лу Ф, Го И, Чжан Джи, Сюй Ф, Ши И, Шен С., Лян Дж. И Гуо В: Аберрант опосредованное метилированием подавление днРНК MEG3 функционирует как aceRNA при раке пищевода.Mol Cancer Res. 15: 800–810. 2017. Просмотр статьи: Google Scholar: PubMed / NCBI | |
16 | Ван К.К. и Чанг Х.Й .: Молекулярные механизмы длинных некодирующих РНК. Mol Cell. 43: 904–914. 2011. Просмотр статьи: Google Scholar: PubMed / NCBI | |
17 | Салмена Л., Полисено Л., Тай Й, Кац Л. и Пандольфи П.П.: Гипотеза цРНК: Розеттский камень скрытой РНК язык? Клетка. 146: 353–358.2011. Просмотр статьи: Google Scholar: PubMed / NCBI | |
18 | Льюис Б.П., Бердж CB и Бартель Д.П .: Консервативное соединение семян, часто окруженное аденозинами, указывает на то, что тысячи генов человека являются мишенями для микроРНК. Клетка. 120: 15–20. 2005. Просмотр статьи: Google Scholar: PubMed / NCBI | |
19 | Зейтц Х: Новое определение мишеней микроРНК. Curr Биол. 19: 870–873. 2009. Просмотр статьи: Google Scholar: PubMed / NCBI | |
20 | Го Г, Кан Цюй, Чжу Х, Чен Цюй, Ван Х, Чен Y, Ouyang J, Zhang L, Tan H, Chen R и др.: Длинная некодирующая РНК критически регулирует клеточную трансформацию, опосредованную Bcr-Abl, посредством выступая в качестве конкурентной эндогенной РНК.Онкоген. 34: 1768–1779. 2015. Просмотр статьи: Google Scholar: PubMed / NCBI | |
21 | Ergun S и Oztuzcu S: Онкозаболевания: CeRNA-опосредованный перекрестный обмен посредством размягчения miRNA в онкогенных путях. Tumor Biol. 36: 3129–3136. 2015. Просмотр статьи: Google Scholar: PubMed / NCBI | |
22 | Джалали С., Бхартия Д., Лалвани М.К., Sivasubbu S и Scaria V: систематический анализ транскриптома Взаимодействия днРНК-миРНК.PLoS One. 8: e538232013. Просмотр статьи: Google Scholar: PubMed / NCBI | |
23 | Люй М.Х., Тан Б., Цзэн С., Ху СиДжей, Се Р., Ву YY, Wang SM, He FT и Yang SM: длинная некодирующая РНК BC032469, a новая конкурирующая эндогенная РНК, усиливает экспрессию hTERT с помощью губка miR-1207-5p и способствует пролиферации при раке желудка. Онкоген. 35: 3524–3534. 2016. Просмотреть статью: Google Scholar: PubMed / NCBI | |
24 | Karreth FA и Pandolfi PP: ceRNA перекрестный разговор при раке: когда соперничество между се-блингами идет наперекосяк.Рак Discov. 3: 1113–1121. 2013. Просмотр статьи: Google Scholar: PubMed / NCBI | |
25 | Гуо Л.Л., Сон СН, Ван П, Дай ЛП, Чжан Джи и Ван К.Дж.: Конкурирующие сети эндогенных РНК и рак желудка. Мир Дж. Гастроэнтерол. 21: 11680–11687. 2015. Просмотр статьи: Google Scholar: PubMed / NCBI | |
26 | Ван И, Сюй Цзинь, Цзян Дж, Сюй Ц, Кан Дж, Сяо L, Wu M, Xiong J, Guo X и Liu H: губка эндогенной miRNA. lincRNA-RoR регулирует Oct4, Nanog и Sox2 в эмбриональном стволе человека самообновление клеток.Dev Cell. 25: 69–80. 2013. Просмотр статьи: Google Scholar: PubMed / NCBI | |
27 | Ørom UA и Shiekhattar R: Длинное некодирование РНК и энхансеры. Curr Opin Genet Dev. 21: 194–198. 2011 г. Просмотр статьи: Google Scholar: PubMed / NCBI | |
28 | Кенири А., Оксли Д., Монье П., Киба М., Дандоло Л., Смитс Дж. И Рейк В.: lincRNA h29 является развивающимся резервуар miR-675, который подавляет рост и Igf1r.Nat Cell Биол. 14: 659–665. 2012. Просмотр статьи: Google Scholar: PubMed / NCBI | |
29 | Да Сакко Л. и Масотти А. Последние выводы и новые инструменты биоинформатики для понимания роли микроРНК связывание с 5′-нетранслируемой областью. Int J Mol Sci. 14: 480–495. 2012 г. Просмотр статьи: Google Scholar: PubMed / NCBI | |
30 | Лопес-Уррутия Э., Бустаманте Монтес LP, Ладрон де Гевара Сервантес D, Перес-Пласенсиа C и Кампос-Парра AD: перекрестные помехи между длинными некодирующими РНК, микро-РНК и мРНК: Расшифровка молекулярных механизмов главных регуляторов рака.Фасад Онкол. 9: 66 | . Просмотр статьи: Google Scholar: PubMed / NCBI |
31 | Го Л., Чжао Ю., Ян С., Чжан Х., Ву Ц. и Чен Ф .: Комплексный эволюционный анализ miRNA-lncRNA в млекопитающие. Mol Biol Rep. 41: 201–207. 2014. Просмотреть статью: Google Scholar: PubMed / NCBI | |
32 | Чжан З., Сунь Л., Чжан И, Лу Г, Ли И и Wei Z: длинная некодирующая РНК FEZF1-AS1 способствует раку груди стволовость и туморогенез посредством нацеливания на ось miR-30a / Nanog.J Cell Physiol. 233: 8630–8638. 2018. Просмотр статьи: Google Scholar: PubMed / NCBI | |
33 | Чжан Л., Ян Ф, Юань Дж. Х., Юань С. X., Чжоу WP, Huo XS, Xu D, Bi HS, Wang F и Sun SH: эпигенетическая активация семейства MiR-200 способствует опосредованному h29 метастазированию подавление гепатоцеллюлярной карциномы. Канцерогенез. 34: 577–586. 2013. Просмотр статьи: Google Scholar: PubMed / NCBI | |
34 | Sun M и Kraus W: От открытия до функция: возрастающая роль длинных некодирующих РНК в физиологии. и болезнь.Endocr Rev.7: er0000999 | . (EPUB перед печатью). Просмотр статьи: Google Scholar |
35 | Юань Дж.Х., Ян Ф., Чен Б.Ф., Лу З., Хо Х.С., Zhou WP, Wang F и Sun SH: гистоновая деацетилаза 4 / SP1 / microrna-200a регулирующая сеть способствует отклонению ацетилирование гистонов при гепатоцеллюлярной карциноме. Гепатология. 54: 2025–2035. 2011. Просмотр статьи: Google Scholar: PubMed / NCBI | |
36 | Цао П, Дэн З., Ван М., Хуанг В., Крамер С.Д., Xu J, Lei M и Sui G: MicroRNA-101 отрицательно регулирует Ezh3 и его экспрессия модулируется рецептором андрогенов и HIF-1альфа / HIF-1бета.Молочный рак. 9: 1082010. Просмотр статьи: Google Scholar: PubMed / NCBI | |
37 | Сандер С., Буллингер Л., Клаппрот К., Фидлер К., Кестлер Х.А., Барт Т.Ф., Мёллер П., Стилгенбауэр С., Поллак JR и Wirth T: MYC стимулирует экспрессию EZh3 путем репрессии его отрицательный регулятор miR-26a. Кровь. 112: 4202–4212. 2008. Просмотр статьи: Google Scholar: PubMed / NCBI | |
38 | Benetatos L, Voulgaris E, Vartholomatos G и Hatzimichael E: Некодирующие РНК и взаимодействия EZh3 в рак: Длинные и короткие рассказы из транскриптома.Int J Cancer. 133: 267–274. 2013. Просмотр статьи: Google Scholar: PubMed / NCBI | |
39 | Лу И, Чжао Х, Лю Цюй, Ли Ц, Грейвс-Дил Р., Cao Z, Singh B, Franklin JL, Wang J, Hu H и др.: LncRNA Произведенные из MIR100HG miR-100 и miR-125b опосредуют устойчивость к цетуксимабу через передачу сигналов Wnt / β-catenin. Nat Med. 23: 1331–1341. 2017 г. Просмотр статьи: Google Scholar: PubMed / NCBI | |
40 | Эрдманн В.А., Шимански М., Хохберг А., Грут N и Barciszewski J: База данных некодирующих мРНК-подобных РНК Y2K.Nucleic Acids Res. 28: 197–200. 2000. Просмотр статьи: Google Scholar: PubMed / NCBI | |
41 | Митуяма Т., Ямада К., Хаттори Э., Окида Х., Оно Й, Тераи Дж., Йошизава А., Комори Т. и Асаи К.: Функциональный RNA Database 3.0: базы данных для поддержки интеллектуального анализа и аннотации функциональные РНК. Nucleic Acids Res. 37: D89 – D92. 2009. Просмотр статьи: Google Scholar: PubMed / NCBI | |
42 | Динджер М.Э., Панг К.С., Мерсер Т.Р., Кроу М.Л., Grimmond SM и Mattick JS: NRED: база данных длинных некодирующих РНК выражение.Nucleic Acids Res. 37: D122 – D126. 2009. Просмотр статьи: Google Scholar: PubMed / NCBI | |
43 | Амарал П.П., Кларк МБ, Гаскойн Д.К., Дингер ME и Mattick JS: lncRNAdb: давно справочная база данных некодирующие РНК. Nucleic Acids Res. 39: D146 – D151. 2011. Просмотр статьи: Google Scholar: PubMed / NCBI | |
44 | Ян Дж. Х., Ли Дж. Х., Цзян С., Чжоу Х. и Цюй Л. Х .: ChIPBase: база данных для расшифровки регуляции транскрипции длинные некодирующие гены РНК и микроРНК из данных ChIP-Seq.Нуклеиновая Acids Res. 41: D177 – D187. 2013. Просмотр статьи: Google Scholar: PubMed / NCBI | |
45 | Volders PJ, Helsens K, Wang X, Menten B, Martens L, Gevaert K, Vandesompele J и Mestdagh P: LN Cipedia: A база данных аннотированных последовательностей транскриптов днРНК человека и конструкции. Nucleic Acids Res. 41: D246 – D251. 2013. Просмотр статьи: Google Scholar: PubMed / NCBI | |
46 | Параскевопулу, доктор медицины, Георгакилас Г, Костулас Н., Рецко М., Марагкакис М., Даламагас TM и Хацигеоргиу AG: DIANA-LncBase: экспериментально подтверждено и вычислено предсказал мишени микроРНК на длинных некодирующих РНК.Нуклеиновые кислоты Res. 41: D239 – D245. 2013. Просмотр статьи: Google Scholar: PubMed / NCBI | |
47 | Повар КБ, Казань Х, Зубери К., Моррис К. и Hughes TR: RBPDB: база данных о специфичности связывания РНК. Нуклеиновая Acids Res. 39: D301 – D308. 2011. Просмотр статьи: Google Scholar: PubMed / NCBI | |
48 | Ли Дж.Х., Лю С., Чжоу Х., Цюй Л.Х. и Ян Дж.Х .: starBase v2.0: декодирование miRNA-ceRNA, miRNA-ncRNA и белок-РНК сети взаимодействия из крупномасштабных данных CLIP-Seq.Нуклеиновые кислоты Res. 42: D92 – D97. 2014. Просмотреть статью: Google Scholar: PubMed / NCBI | |
49 | Hsu PW, Huang HD, Hsu SD, Lin LZ, Tsou AP, Tseng CP, Stadler PF, Washietl S и Hofacker IL: miRNAMap: Genomic карты генов микроРНК и их генов-мишеней в геномах млекопитающих. Nucleic Acids Res. 34: D135 – D139. 2006. Просмотр статьи: Google Scholar: PubMed / NCBI | |
50 | Цуй Т, Чжан Л., Хуан И, И Й, Тан П, Чжао Y, Hu Y, Xu L, Li E и Wang D: MNDR v2.0: Обновленный ресурс Ассоциации нкРНК-болезнь у млекопитающих. Nucleic Acids Res. 46: D371 – D374. 2018.PubMed / NCBI | |
51 | Ян Б., Ван Чж и Го Дж. Т.: Исследование стратегии для исследования функции длинных некодирующих РНК. Геномика. 99: 76–80. 2012. Просмотр статьи: Google Scholar: PubMed / NCBI | |
52 | Ørom UA, Дерриен Т., Берингер М., Гумиредди К, Гардини А., Буссотти Дж., Лай Ф., Зитницки М., Нотредам С, Хуанг К., и др.: Длинные некодирующие РНК с энхансер-подобной функцией у человека клетки.Клетка. 143: 46–58. 2010. Просмотр статьи: Google Scholar: PubMed / NCBI | |
53 | Лин М, Педроса Э, Шах А, Грабовский А, Maqbool S, Zheng D и Lachman HM: последовательность РНК человеческих нейронов из iPS-клеток выявляет кандидатные длинные некодирующие РНК, участвующие в нейрогенез и нервно-психические расстройства. PLoS One. 6: e233562011. Просмотр статьи: Google Scholar: PubMed / NCBI | |
54 | Алвин Дж. К., Кемп Диджей и Старк Г. Р.: Метод для обнаружения специфических РНК в агарозных гелях путем переноса в диазобензилоксиметилбумага и гибридизация с ДНК-зондами.Proc Natl Acad Sci USA. 74: 5350–5354. 1977 г. Просмотреть статью: Google Scholar: PubMed / NCBI | |
55 | Хайд Калифорния, Стивенс Дж., Ливак К.Дж. и Уильямс PM: Количественная ПЦР в реальном времени. Genome Res. 6: 986–994. 1996 г. Просмотр статьи: Google Scholar: PubMed / NCBI | |
56 | Чакраборти Д., Каппей Д., Тайс М., Ницше А, Динг Л., Пашковски-Рогач М., Сурендранат В., Бергер Н., Шульц Х., Саар К. и др.: Комбинированная РНКи и локализация функционально рассечение длинных некодирующих РНК.Нат методы. 9: 360–362. 2012 г. Просмотр статьи: Google Scholar: PubMed / NCBI | |
57 | Альбертс Б., Джонсон А. и Льюис Дж.: Фракционирование клеток. Молекулярная биология клетки. 4-й. Новый Йорк: Наука о гирляндах; 2002 | |
58 | Кошкин А.А., Сингх С.К., Нильсен П., Раджванши VK, Kumar R, Meldgaard M, Olsen CE и Wengel J: LNA (закрыто Нуклеиновые кислоты): синтез аденина, цитозина, гуанина, 5-метилцитозин, тимин и мономеры бициклонуклеозидов урацила, олигомеризация и беспрецедентное распознавание нуклеиновых кислот.Тетраэдр. 54: 3607–3630. 1998. Просмотр статьи: Google Scholar | |
59 | Чжан Ф., Вэнь И и Го Х: CRISPR / Cas9 для редактирование генома: прогресс, последствия и проблемы. Хум Мол Genet. 23: R40 – R46. 2014. Просмотреть статью: Google Scholar: PubMed / NCBI | |
60 | Огонь А, Сюй С., Монтгомери М.К., Костас С.А., Driver SE и Mello CC: сильное и специфическое генетическое вмешательство со стороны двухцепочечная РНК у Caenorhabditis elegans.Природа. 391: 806–811. 1998. Просмотр статьи: Google Scholar: PubMed / NCBI | |
61 | Кин Дж. Д., Комисаров Дж. М. и Фридерсдорф MB: RIP-Chip: выделение и идентификация мРНК, микроРНК и белковые компоненты рибонуклеопротеидных комплексов из клетки экстракты. Nat Protoc. 1: 302–307. 2006. Просмотр статьи: Google Scholar: PubMed / NCBI | |
62 | Чжао Дж., Осуми Т.К., Кунг Дж. Т., Огава Й., Грау DJ, Сарма К., Сон Джей Джей, Kingston RE, Borowsky M и Lee JT: Полногеномная идентификация поликомб-ассоциированных РНК с помощью RIP-seq.Mol Cell. 40: 939–953. 2010. Просмотр статьи: Google Scholar: PubMed / NCBI | |
63 | Беллуччи М, Агостини Ф, Масин М и Tartaglia GG: прогнозирование белковых ассоциаций с длительным некодированием РНК. Нат методы. 8: 444–445. 2011. Просмотр статьи: Google Scholar: PubMed / NCBI | |
64 | Lagarde J, Uszczynska-Ratajczak B, Carbonell S, Pérez-Lluch S, Abad A, Davis C, Gingeras TR, Frankish A, Харроу Дж., Гиго Р. и Джонсон Р.: высокопроизводительная аннотация полноразмерные длинные некодирующие РНК с захватом долгого чтения.Нат Жене. 49: 1731–1740. 2017. Просмотр статьи: Google Scholar: PubMed / NCBI | |
65 | Танизава Х. и Нома К. организация генома с помощью 3C-seq. Semin Cell Dev Biol. 23: 213–221. 2012. Просмотр статьи: Google Scholar: PubMed / NCBI | |
66 | Сплинтер Э, де Вит Э, ван де Веркен Х. Дж., Клоус П. и де Лаат В. Определение дальнего хроматина взаимодействия для выбранных геномных сайтов с использованием технологии 4C-seq: От фиксации к вычислению.Методы. 58: 221–230. 2012. Просмотр статьи: Google Scholar: PubMed / NCBI | |
67 | Дости Дж. И Деккер Дж.: Отображение сетей физические взаимодействия между элементами генома с использованием технологии 5C. Nat Protoc. 2: 988–1002. 2007. Просмотр статьи: Google Scholar: PubMed / NCBI | |
68 | Белтон Дж. М., МакКорд Р. П., Гибкус Дж. Х., Наумова N, Zhan Y и Dekker J: Hi-C: комплексная техника захвата конформация геномов.Методы. 58: 268–276. 2012. Просмотр статьи: Google Scholar: PubMed / NCBI | |
69 | Северс, штат Нью-Джерси: замораживание-разрушение электронов микроскопия. Nat Protoc. 2: 547–576. 2007. Просмотр статьи: Google Scholar: PubMed / NCBI | |
70 | Оливари С., Плесси С. и Карнинчи П.: Высокопроизводительная проверка сайтов начала транскрипции с помощью Глубокая гонка. Биотехнологии. 46: 130–132. 2009. Просмотр статьи: Google Scholar: PubMed / NCBI | |
71 | Cossu AM, Mosca L, Zappavigna S, Misso G, Bocchetti M, De Micco F, Quagliuolo L, Porcelli M, Caraglia M и Boccellino M: длинные некодирующие РНК как важные биомаркеры в рак гортани и другие опухоли головы и шеи.Int J Mol Sci. 20: E34442019. Просмотр статьи: Google Scholar: PubMed / NCBI | |
72 | Наппи L и Николс C: МикроРНК как биомаркеры половых опухолей. Urol Clin North Am. 46: 449–457. 2019. Просмотр статьи: Google Scholar: PubMed / NCBI | |
73 | Цзян Ю.Ф., Чжан Х.Й., Ке Дж., Шен Х., Оу HB и Liu Y: сверхэкспрессия LncRNA GHET1 предсказывает неблагоприятный выживаемость и клинические параметры пациентов с различными видами рака.J Cell Mol Med. 23: 4891–4899. 2019. Просмотр статьи: Google Scholar: PubMed / NCBI | |
74 | Moody L, Dvoretskiy S, An R, Mantha S и Pan YX: эффективность miR-20a как диагностического и прогностического биомаркер колоректального рака: систематический обзор и метаанализ. Раки (Базель). 11: E11112019. Просмотр статьи: Google Scholar: PubMed / NCBI | |
75 | Пермут Дж. Б., Чен Д. Т., Йодер С. Дж., Ли Дж., Смит AT, Чой Дж. У., Ким Дж., Балагурунатан Й., Цзян К., Коппола Д. и др.: Использование циркулирующих длинных некодирующих РНК для диагностики и Злокачественное прогнозирование внутрипротоковых папиллярных муцинозных новообразований Поджелудочная железа.Sci Rep.7: 104842017. Просмотр статьи: Google Scholar: PubMed / NCBI | |
76 | Ли А., Ю Дж, Ким Х, Вольфганг К.Л., Канто М.И., Hruban RH и Goggins M: сыворотка miR-1290 как маркер поджелудочной железы. рак-ответ. Clin Cancer Res. 19: 5252–5253. 2013. Просмотр статьи: Google Scholar: PubMed / NCBI | |
77 | Ян Цюй, Тан И, Тан Ц, Цун Ч, Ван Х, Shen X и Ju S: снижение экспрессии LINC00173, индуцированной miR-182-5p накопление способствует пролиферации, миграции и апоптозу клеток ингибирование через путь AGER / NF-κB при немелкоклеточном раке легкого.Являюсь J Transl Res. 11: 4248–4262. 2019.PubMed / NCBI | |
78 | Ли XY, Чжоу LY, Luo H, Zhu Q, Zuo L, Liu GY, Feng C, Zhao JY, Zhang YY и Li X: длинная некодирующая РНК MIR210HG способствует метастазированию опухоли, действуя как цеРНК miR-1226-3p для регуляции экспрессии муцина-1c в инвазивной груди рак. Старение (Олбани, штат Нью-Йорк). 11: 5646–5665. 2019.PubMed / NCBI | |
79 | Чжан Ю., Чжу З., Хуан С., Чжао Ц., Хуан Ц., Тан Y, Sun C, Zhang Z, Wang L, Chen H и др.: LncRNA XIST регулирует пролиферацию и миграцию гепатоцеллюлярной карциномы клетки, действуя как молекулярная губка miR-497-5p и нацеливаясь на PDCD4.Cancer Cell Int. 19: 1982019. Просмотр статьи: Google Scholar: PubMed / NCBI | |
80 | Ху Л, Е Х, Хуан Г, Ло Ф, Лю И, Лю И, Ян X, Шен Дж, Лю Кью и Чжан Дж .: Длинная некодирующая РНК GAS5 подавляет миграцию и инвазию гепатоцеллюлярной карциномы клетки через miR-21. Tumor Biol. 37: 2691–2702. 2016. Просмотреть статью: Google Scholar: PubMed / NCBI | |
81 | У Х, Чжэн Дж, Дэн Дж, Чжан Л., Ли Н, Ли У, Ли Ф, Лу Дж и Чжоу У: LincRNAuc002yug.2 включает в себя альтернативу сплайсинг RUNX1 и служит предиктором рака пищевода и прогноз. Онкоген. 34: 4723–4734. 2015. Просмотр статьи: Google Scholar: PubMed / NCBI | |
82 | Чэнь Д., Чжан З., Мао Ц., Чжоу Ю., Ю Л, Инь Y, Wu S, Mou X и Zhu Y: ANRIL ингибирует p15 (INK4b) через Сигнальный путь TGFβ1 в плоских клетках пищевода человека карцинома. Cell Immunol. 289: 91–96. 2014. Просмотреть статью: Google Scholar: PubMed / NCBI | |
83 | Кан М., Сан И, Гу Х, Чжэн Л., Ван Л., Лю C, Shi Y, Shao A, Ding G, Chen G и др.: Длинные некодирующие РНК POLR2E rs3787016 C / T и полиморфизмы HULC rs7763881 A / C связаны со сниженным риском рака пищевода.Tumor Biol. 36: 6401–6408. 2015. Просмотр статьи: Google Scholar: PubMed / NCBI | |
84 | Чжан Э, Хан Л, Инь Д, Хэ Х, Хун Л, Си Х, Qiu M, Xu T, De W, Xu L, et al: ацетилирование h4K27, активированное долго некодирующая РНК CCAT1 влияет на пролиферацию и миграцию клеток посредством регулирование экспрессии SPRY4 и HOXB13 в плоскоклеточных клетках пищевода карцинома. Nucleic Acids Res. 45: 3086–3101. 2017. Просмотр статьи: Google Scholar: PubMed / NCBI | |
85 | Маццу Ю.З., Йошикава Ю., Нандакумар С., Чакраборти Дж., Армения Дж., Джехан Л. Э., Ли Дж. М. и Кантофф П. В.: Связанное с метилированием сайленсинг miR-193b активирует главные драйверы агрессивного рака простаты.Мол Онкол. 13: 1944–1958. 2019. Просмотр статьи: Google Scholar: PubMed / NCBI | |
86 | Чен Г, Фэн Й, Ли Х, Цзян З, Бэй Б, Zhang L, Han Y, Li Y и Li N: посттранскрипционная регуляция генов при раке, ассоциированном с колитом. Фронт Жене. 10: 5852019. Просмотр статьи: Google Scholar: PubMed / NCBI | |
87 | Кадир М.И., Бухат С., Расул С., Манзур Х. и Манзур М: РНК-терапия: идентификация новых мишеней ведущие к открытию лекарств.J Cell Biochem. 2019: (Epub опережает Распечатать). | |
88 | Мэн Х, Ли Х, Чжан П, Ван Дж, Чжоу И и Чен М: Круговая РНК: новый ключевой игрок в мире РНК. Краткий Биоинформ. 18: 547–557. 2017.PubMed / NCBI | |
89 | Цюй С, Ян Х, Ли Х, Ван Дж, Гао И, Шан R, Sun W, Dou K и Li H: круговая РНК: новая звезда некодирования РНК. Cancer Lett. 365: 141–148. 2015. Просмотр статьи: Google Scholar: PubMed / NCBI | |
90 | Sato K и Siomi MC: взаимодействие с Piwi РНК: биологические функции и биогенез.Очерки Биохимии. 54: 39–52. 2013. Просмотр статьи: Google Scholar: PubMed / NCBI | |
91 | Буш Дж., Ралла Б., Юнг М., Вотчофски З., Трухильо-Аррибас Э., Швабе П., Килич Э., Фендлер А. и Юнг К.: Piwi-взаимодействующие РНК как новые прогностические маркеры в светлой клетке почечно-клеточный рак. J Exp Clin Cancer Res. 34: 612015. Просмотр статьи: Google Scholar: PubMed / NCBI |
Brunswick Gold Ltd приобретает подразделения Otso Gold Corp.
НИКОСИЯ, Кипр, 9 февраля 2021 г. / CNW / — Этот пресс-релиз выпущен компанией Brunswick Gold Ltd (« Brunswick Gold ») в соответствии с требованиями к раннему предупреждению Национальных инструментов Канады 62-104 и 62- 103 в отношении простых акций Otso Gold Corp. (« Эмитент »).
Brunswick Gold объявляет, что она завершила коммерческое частное размещение ценных бумаг с Эмитентом для стратегических инвестиций в размере 11 миллионов долларов США (« Финансирование »).В соответствии с Финансированием, Brunswick Gold было выпущено 284 944 440 единиц Эмитента (каждая, « единиц ») по цене выпуска, равной 0,05 канадских долларов за единицу. Каждая Пая состояла из одной обыкновенной акции (каждая, « Простая акция ») и одного ордера на покупку Обыкновенной акции (каждая, « Варранта »). Каждый варрант дает Brunswick Gold право на покупку одной обыкновенной акции по цене исполнения, равной 0,05 канадского доллара за варрант, в течение пяти (5) лет с даты выпуска. 25 904 040 Паев, выпущенных Эмитентом для Brunswick Gold, были использованы для погашения основной суммы предыдущего необеспеченного неконвертируемого займа в размере 1 млн долларов США от Brunswick Gold для Эмитента.Финансирование было основано на обменном курсе канадских долларов к доллару США, составляющем 1,29520 канадских долларов, и было одобрено акционерами эмитента на годовом и внеочередном собрании акционеров, состоявшемся 20 января 2021 года.
По завершении финансирования Brunswick Gold приобрела выгодную владение и контроль над 569 888 880 Обыкновенными акциями (в силу его владения и контроля над 284 944 440 Простыми акциями и 284 944 440 Варрантами), что составляет приблизительно 46,03% выпущенных и находящихся в обращении Обыкновенных акций при завершении на неразводненной основе и приблизительно 63.05% выпущенных и находящихся в обращении Простых акций на частично разводненной основе при условии исполнения всех варрантов, бенефициарными собственниками и контролем которой является Brunswick Gold.
Brunswick Gold приобрела Простые акции и Варранты в инвестиционных целях. Как описано в информационном проспекте управления, подготовленном Эмитентом в связи с собранием, и как предусмотрено в соответствии с условиями соглашения о правах инвестора, заключенного между Эмитентом и Brunswick Gold, совет директоров Эмитента был восстановлен после закрытия Финансирования до состоит из следующих семи директоров: Брайан Вессон, Клайд Вессон, Иветт Харрисон и четыре кандидата от Brunswick Gold (Владимир Лелеков, Николя Паско, Виктор Кошкин и Мартин Смит).Кроме того, Председателем Совета директоров назначен Владимир Лелеков. Соглашение о правах инвестора также предоставляет Brunswick Gold право назначать финансового контролера Эмитента и некоторые другие права на обычную регистрацию и преимущественную подписку в отношении будущих продаж ценных бумаг Эмитентом.
История продолжается
Для получения дополнительной информации и для получения копии отчета о раннем предупреждении, поданного в соответствии с применимым канадским провинциальным и территориальным законодательством о ценных бумагах в связи со Сделкой, перейдите в профиль Эмитента на веб-сайте SEDAR (www.sedar.com) или свяжитесь с Андреасом Орфанидесом по телефону [email protected] . Зарегистрированный офис Brunswick Gold находится по адресу 2-4 Arch Makarios III Avenue, Capital Center, 9th Floor, Nicosia 1065, Cyprus.
ИСТОЧНИК Brunswick Gold Ltd
Cision
См. Исходное содержание: http://www.newswire.ca/en/releases/archive/Feb February2021/09/c3638.