УПП — это… Что такое УПП?
УППучебно-производственное предприятие
организация
Словарь: С. Фадеев. Словарь сокращений современного русского языка. — С.-Пб.: Политехника, 1997. — 527 с.
УППустановка для приготовления пульпы
УППустройство постоянной памяти
комп.
УППуправление производственных предприятий
организация
Словарь: С. Фадеев.
Управление процессами перевозок
факульт ЛИИЖТ
образование и наука
УППустановка полевая проверочная
УППустройство плавного пуска
УППуправление пищевой промышленности, упрпищепром
УППучётная политика предприятия
бух., фин.
УППучастковый пункт полиции
УППудалить по плану
Источник: http://www. tehlit.ru/1lib_norma_doc/52/52115/#i48556
tehlit.ru/1lib_norma_doc/52/52115/#i48556
управление промышленных предприятий
например: УПП при Спецстрое России
УППуглеперерабатывающее предприятие
УППуравнение первого порядка
УППдисциплина в вузе
образование и наука, юр.
Источник: http://www.booka.ru/booka_topic_75821?page=5
УППуниверсальная платёжная платформа
Источник: http://gazeta. etatar.ru/news/view/8/27870
etatar.ru/news/view/8/27870
Уральская правовая палата
с 2005
юр.
Источник: http://www.expert.ru/printissues/ural/2006/45/yuridicheskie_uslugi/
УППУправление по подотчётности правительству
англ.: GAO, Government accountability office
англ.: US GAO, U.S. Government accountability office
Счётная палата США
англ., США
Источник: http://www.izvestia.ru/news/news116995/
УПП«Управление производственным предприятием»
программа 1С
УППуправление пассажирских перевозок
Источник:
 regnum.ru/news/548274.html
regnum.ru/news/548274.htmlПример использования
УПП в Новосибирске
УППучебно-производственная практика
Источник: http://www.vfrmat.ru/kafedra_turizma.html
Пример использования
отдел УПП
Словарь сокращений и аббревиатур. Академик. 2015.
УПП — это… Что такое УПП?
УПП — учебно производственное предприятие организация Словарь: С. Фадеев. Словарь сокращений современного русского языка. С. Пб.: Политехника, 1997. 527 с. УПП установка для приготовления пульпы УПП устройство постоянной памяти комп … Словарь сокращений и аббревиатур
УПП
— управление пищевой промышленности управление производственных предприятий установка для приготовления пульпы установка полевая проверочная устройство постоянной памяти (в ЭВМ) учебно производственное предприятие … Словарь сокращений русского языкаУПП ГМС — Управление промышленных предприятий Главмосстроя организация … Словарь сокращений и аббревиатур
УПП КОС — учебно производственное предприятие Казахского общества слепых Казахстан, образование, организация … Словарь сокращений и аббревиатур
УПП «Вектор» — ОАО «Уральское производственное предприятие «Вектор» военное предприятие, специализирующееся на производстве аппаратуры связи, товаров народного потребления и производственно технического назначения. Располагается в Екатеринбурге. Входит в состав … Википедия
Располагается в Екатеринбурге. Входит в состав … Википедия
Код УПП
Мичуринск УПП — 393730, Тамбовской, Мичуринского … Населённые пункты и индексы России
Лимит — (Limit) Содержание Содержание Определения описываемого предмета Лимитирование банковских операций Позиционные Объемные лимиты Лимиты на характеристики позиций, на взвешенный объем Структурные лимиты (долевые лимиты, лимиты концентрации) Лимиты… … Энциклопедия инвестора
Устройство плавного пуска — Простейшее электрическое устройство плавного пуска двигателя постоянного тока Устройство плавного пуска электротехническое(электронное) устройство, используемое для плавного пуска асинхронных электродвигателей с … Википедия
Днепропетровское учебно-производственное предприятие Украинского общества слепых — Днепропетровское УПП УТОС Год основания 1952 Расположение … Википедия
Устройство плавного пуска электродвигателя.
 Как это работает.
Как это работает.Устройство плавного пуска — электротехническое устройство, используемое в асинхронных электродвигателях, которое позволяет во время запуска удерживать параметры двигателя (тока, напряжения и т.д.) в в безопасных пределах. Его применение уменьшает пусковые токи, снижает вероятность перегрева двигателя, устраняет рывки в механических приводах, что, в конечном итоге, повышает срок службы электродвигателя.
НазначениеУправление процессом запуска, работы и остановки электродвигателей. Основными проблемами асинхронных электродвигателей являются:
- невозможность согласования крутящего момента двигателя с моментом нагрузки,
- высокий пусковой ток.
Во время пуска крутящий момент за доли секунды часто достигает 150-200%, что может привести к выходу из строя кинематической цепи привода. При этом стартовый ток может быть в 6-8 раз больше номинального, порождая проблемы со стабильностью питания.
Основной проблемой асинхронных электродвигателей является то, что момент силы, развиваемый электродвигателем, пропорционален квадрату приложенного к нему напряжения, что создаёт резкие рывки ротора при пуске и остановке двигателя, которые, в свою очередь, вызывают большой индукционный ток.
Софтстартеры могут быть как механическими, так и электрическими, либо сочетать то и другое.
Механические устройства непосредственно противодействуют резкому нарастанию оборотов двигателя, ограничивая крутящий момент. Они могут представлять собой тормозные колодки, жидкостные муфты, магнитные блокираторы, противовесы с дробью и прочее.
Данные электрические устройства позволяют постепенно повышать ток или напряжение от начального пониженного уровня (опорного напряжения) до максимального, чтобы плавно запустить и разогнать электродвигатель до его номинальных оборотов. Такие УПП обычно используют амплитудные методы управления и поэтому справляются с запуском оборудования в холостом или слабо нагруженном режиме. Более современное поколение УПП (например, устройства ЭнерджиСейвер) используют фазовые методы управления и потому способны запускать электроприводы, характеризующиеся тяжелыми пусковыми режимами «номинал в номинал». Такие УПП позволяют производить запуски чаще и имеют встроенный режим энергосбережения и коррекции коэффициента мощности.
Выбор устройства плавного пуска
При включении асинхронного двигателя в его роторе на короткое время возникает ток короткого замыкания, сила которого после набора оборотов снижается до номинального значения, соответствующего потребляемой электрической машиной мощности. Это явление усугубляется тем, что в момент разгона скачкообразно растет и крутящий момент на валу. В результате может произойти срабатывание защитных автоматических выключателей, а если они не установлены, то и выход из строя других электротехнических устройств, подключенных к той же линии. И в любом случае, даже если аварии не произошло, при пуске электромоторов отмечается повышенный расход электроэнергии. Для компенсации или полного устранения этого явления используются устройства плавного пуска (УПП).
Это явление усугубляется тем, что в момент разгона скачкообразно растет и крутящий момент на валу. В результате может произойти срабатывание защитных автоматических выключателей, а если они не установлены, то и выход из строя других электротехнических устройств, подключенных к той же линии. И в любом случае, даже если аварии не произошло, при пуске электромоторов отмечается повышенный расход электроэнергии. Для компенсации или полного устранения этого явления используются устройства плавного пуска (УПП).
Как реализуется плавный пуск
Чтобы плавно запустить электродвигатель и не допустить броска тока, используются два способа:
- Ограничивают ток в обмотке ротора. Для этого ее делают состоящей из трех катушек, соединенных по схеме «звезда». Их свободные концы выводят на контактные кольца (коллекторы), закрепленные на хвостовике вала. К коллектору подключают реостат, сопротивление которого в момент пуска максимальное. По мере его снижения ток ротора растет и двигатель раскручивается.
 Такие машины называются двигателями с фазным ротором. Они используются в крановом оборудовании и в качестве тяговых электромоторов троллейбусов, трамваев.
Такие машины называются двигателями с фазным ротором. Они используются в крановом оборудовании и в качестве тяговых электромоторов троллейбусов, трамваев. - Уменьшают напряжение и токи, подаваемые на статор. В свою очередь, это реализуется с помощью:
а) автотрансформатора или реостата;
б) ключевыми схемами на базе тиристоров или симисторов.
Именно ключевые схемы и являются основой построения электротехнических приборов, которые принято назвать устройствами плавного пуска или софтстартерами. Обратите внимание, что частотные преобразователи так же позволяют плавно запустить электродвигатель, но они лишь компенсируют резкое возрастание крутящего момента, не ограничивая при этом пускового тока.
Принцип работы ключевой схемы основывается на том, что тиристоры отпираются на определенное время в момент прохождения синусоидой ноля. Обычно в той части фазы, когда напряжение растет. Реже – при его падении. В результате на выходе УПП регистрируется пульсирующее напряжение, форма которого лишь приблизительно похожа на синусоиду. Амплитуда этой кривой растет по мере того, как увеличивается временной интервал, когда тиристор отперт.
Реже – при его падении. В результате на выходе УПП регистрируется пульсирующее напряжение, форма которого лишь приблизительно похожа на синусоиду. Амплитуда этой кривой растет по мере того, как увеличивается временной интервал, когда тиристор отперт.
Критерии выбора софтстартера
По степени снижения степени важности критерии выбора устройства располагаются в следующей последовательности:
- Мощность.
- Количество управляемых фаз.
- Обратная связь.
- Функциональность.
- Способ управления.
- Дополнительные возможности.
Мощность
Главным параметром УПП является величина Iном – сила тока, на которую рассчитаны тиристоры. Она должна быть в несколько раз больше значения силы тока, проходящего через обмотку двигателя, вышедшего на номинальные обороты. Кратность зависит от тяжести пуска. Если он легкий – металлорежущие станки, вентиляторы, насосы, то пусковой ток в три раза выше номинального. Тяжелый пуск характерен для приводов, имеющих значительный момент инерции. Таковы, например, вертикальные конвейеры, пилорамы, прессы. Ток выше номинального в пять раз. Существует и особо тяжелый пуск, который сопровождает работу поршневых насосов, центрифуг, ленточных пил… Тогда Iном софтстартера должен быть в 8-10 раз больше.
Тяжелый пуск характерен для приводов, имеющих значительный момент инерции. Таковы, например, вертикальные конвейеры, пилорамы, прессы. Ток выше номинального в пять раз. Существует и особо тяжелый пуск, который сопровождает работу поршневых насосов, центрифуг, ленточных пил… Тогда Iном софтстартера должен быть в 8-10 раз больше.
Тяжесть пуска влияет и на время его завершения. Он может длиться от десяти до сорока секунд. За это время тиристоры сильно нагреваются, поскольку рассеивают часть электрической мощности. Для повторения им надо остыть, а на это уходит столько же, сколько на рабочий цикл. Поэтому если технологический процесс требует частого включения-выключения, то выбирайте софтстартер как для тяжелого пуска. Даже если ваше устройство не нагружено и легко набирает обороты.
Количество фаз
Можно управлять одной, двумя или тремя фазами. В первом случае устройство в большей степени смягчает рост пускового момента, чем тока. Чаще всего используются двухфазные пускатели. А для случаев тяжелого и особо тяжелого пуска – трехфазные.
Чаще всего используются двухфазные пускатели. А для случаев тяжелого и особо тяжелого пуска – трехфазные.
Обратная связь
УПП может работать по заданной программе – увеличить напряжение до номинала за указанное время. Это наиболее простое и распространенное решение. Наличие обратной связи делает процесс управления более гибким. Параметрами для нее служат сравнение напряжения и вращающего момента или фазный сдвиг между токами ротора и статора.
Функциональность
Возможность работать на разгон или торможение. Наличие дополнительного контактора, который шунтирует ключевую схему и позволяет ей остыть, а также ликвидирует несимметричность фаз из-за нарушения формы синусоиды, которое приводит к перегреву обмоток.
Способ управления
Бывает аналоговым, посредством вращения потенциометров на панели, и цифровым, с применением цифрового микроконтроллера.
Дополнительные функции
Все виды защиты, режим экономии электроэнергии, возможность пуска с рывка, работы на пониженной скорости (псевдочастотное регулирование).
Правильно подобранный УПП увеличивает вдвое рабочий ресурс электродвигателей, экономит до 30 процентов электроэнергии.
Зачем нужно устройство плавного пуска (софтстартера)
Все чаще при запуске электроприводов насосов, вентиляторов применяются устройство плавного пуска (софтстартер). С чем это связано? В нашей статье мы постараемся осветить этот вопрос.
Асинхронные двигатели используются уже более ста лет, и за это время относительно мало изменилось их функционирование. Запуск этих устройств и связанные с ним проблемы хорошо известны их владельцам. Пусковые токи приводят к просадкам напряжения и перегрузкам проводки, вследствие чего:
— некоторая электротехника может самопроизвольно отключаться;
— возможен сбой оборудования и т. д.
д.
Своевременно установленный приобретенный и подключенный софтстартер позволяет избежать лишних трат денег и головной боли.
Что такое пусковой ток
В основе принципа действия асинхронных двигателей лежит явление электромагнитной индукции. Наращивание обратной электродвижущей силы (э. д. с), которая создается путем применения изменяющегося магнитного поля во время запуска двигателя, приводит к переходным процессам в электрической системе. Этот переходной режим может повлиять на систему электропитания и другое оборудование, подключенное к нему.
Во время запуска электродвигатель разгоняется до полной скорости. Продолжительность начальных переходных процессов зависит от конструкции агрегата и характеристик нагрузки. Пусковой момент должен быть наибольшим, а пусковые токи – наименьшими. Последние влекут за собой пагубные последствия для самого агрегата, системы электроснабжения и оборудования, подключенного к нему.
В течение начального периода пусковой ток может достигать пяти-восьмикратного тока полной нагрузки. Во время пуска электродвигателя кабели вынуждены пропускать больше тока, чем во время периода стабильного состояния. Падение напряжения в системе также будет намного больше при пуске, чем во время стабильной работы – это становится особенно очевидным при запуске мощного агрегата или большого числа электродвигателей одновременно.
Способы защиты электродвигателя
Поскольку использование электродвигателей стало широко распространенным, преодоление проблем с их запуском стало проблемой. На протяжении многих лет для решения этих задач были разработано несколько методов, каждый из которых имеет свои преимущества и ограничения.
В последнее время были достигнуты значительные успехи в использовании электроники в регулировании электроэнергии для двигателей. Все чаще при запуске электроприводов насосов, вентиляторов применяются устройство плавного пуска. Всё дело в том, что прибор имеет ряд особенностей.
Всё дело в том, что прибор имеет ряд особенностей.
Особенностью устройства пуска является то, что он плавно подаёт на обмотки двигателя напряжение от нуля до номинального значения, позволяя двигателю плавно разгоняться до максимальной скорости. Развиваемый электродвигателем механический момент пропорционален квадрату приложенного к нему напряжения.
В процессе пуска УПП постепенно увеличивает подаваемое напряжение, и электромотор разгоняется до номинальной скорости вращения без большого момента и пиковых скачков тока.
Виды устройств плавного пуска
На сегодняшний день для плавного запуска техники используются три типа УПП: с одной, двумя и со всеми управляемыми фазами.
Первый тип применяется для однофазного двигателя для обеспечения надежной защиты от перегрузки, перегрева и снижения влияния электромагнитных помех.
Как правило, схема второго типа помимо полупроводниковой платы управления включает в себя байпасный контактор. После того как двигатель раскрутится до номинальной скорости, байпасный контактор срабатывает и обеспечивает прямую подачу напряжения на электродвигатель.
После того как двигатель раскрутится до номинальной скорости, байпасный контактор срабатывает и обеспечивает прямую подачу напряжения на электродвигатель.
Трехфазный тип является самым оптимальным и технически совершенным решением. Он обеспечивает ограничение тока и силы магнитного поля без перекосов по фазам.
Зачем же нужно устройство плавного пуска?
Благодаря относительно невысокой цене популярность софтстартеров набирает обороты на современном рынке промышленной и бытовой техники. УПП для асинхронного электродвигателя необходимо для продления его срока службы. Большим преимуществом софтстартера является то, что пуск осуществляется с плавным ускорением, без рывков.
Есть отличная альтернатива устройству плавного пуска. Стоимость отличается, но и функциональные возможности расширенные.
Преобразователь частоты – это решение задачи, когда требуется регулирование скорости электродвигателя и автоматизация работы технологичного оборудования через обратную связь посредством датчика. При помощи преобразователя Вы сможете решить более сложные и разносторонние вопросы по автоматизации электропривода.
При помощи преобразователя Вы сможете решить более сложные и разносторонние вопросы по автоматизации электропривода.
Остались вопросы?
Специалисты ЭНЕРГОПУСК ответят на Ваши вопросы:
8-800-700-11-54 (8-18, Пн-Вт)
Устойчивость к противомикробным препаратам | Продовольственная и сельскохозяйственная организация Объединенных Наций
Мы разрабатываем новую версию веб-сайта, посвященного вопросам УПП, которая будет в большей степени отвечать потребностям пользователей. В связи с этим мы собираем информацию о том, кто и как пользуется данным веб-сайтом, и предложения по его улучшению. Мы высоко ценим ваши замечания и предложения и учтем их при разработке новой версии веб-сайта. Просьба не позднее 31 мая 2021 года пройти этот опрос и распространить его среди своих коллег и в рамках соответствующих профессиональных сетей. Чтобы ответить на все вопросы, вам понадобится около 14 минут. Сделать это можно анонимно. Заранее благодарим вас за участие в опросе. |
Противомикробным препаратам принадлежит критически важная роль в лечении заболеваний, их применение необходимо для охраны здоровья как человека, так и животных. Однако зачастую противомикробные препараты неправильно используются для лечения и профилактики заболеваний в животноводческом секторе, аквакультуре и растениеводстве. Такие действия нередко связаны с потенциальным риском появления и распространения микроорганизмов, устойчивых к противомикробным препаратам.
Ежегодно от причин, связанных с резистентностью к противомикробным препаратам, в мире умирает порядка 700 тысяч человек (Комиссия Великобритании — О’Нейл) | В ветеринарии используется 27 различных классов противомикробных препаратов | В 2011 году объем глобального рынка ветеринарных препаратов оценивался в 22 млрд долларов США (ОЭСР) | В период с 2015 по 2017 годы количественные данные о использовании противомикробных препаратов в животноводстве представили 118 стран, в то время как в 2015 году такие отчеты представили 89 стран |
 subcl div.csc-default»).append(»)
})
subcl div.csc-default»).append(»)
})Преобразователь частоты термины, устройство мягкого пуска
Преобразователь частоты (ПЧ) — это электронное устройство для изменения частоты электрического тока (напряжения):
- Машина, преобразующая переменной ток одной частоты в переменный ток другой частоты (по СТ МЭК 50(411)-73)
- Преобразователь электрической энергии, который преобразует электрическую мощность с изменением частоты (по СТ МЭК 50(151)-78)
Низковольтный ПЧ – частотный преобразователь, работающий на напряжении 110, 220, 380 или 660 В. Низковольтные преобразователь частоты применяются для управления низковольтными асинхронным и синхронными электродвигателями.
Высоковольтный ПЧ (средневольтный ПЧ) – преобразователь частоты, работающий на напряжениях от 2,1 до 10 кВ. Применяются для управления высоковольтными синхронными и асинхронным электродвигателем.
В различный источниках преобразователь частоты может называться также:
- Частотный преобразователь
- Frequency converter
- Инвертор
- DC/AC Converter
- DC/AC Drive
- Инвертор напряжения
- Инвертор тока
- Частотник
- Электропреобразователь частоты
- Частотно-регулируемый привод
- Тиристорный регулятор напряжения
Frequency converter — частотный преобразователь;
Инвертор — устройство для преобразования постоянного тока в переменный с изменением величины частоты и/или напряжения.
Инвертор напряжения – это инвертор, форма напряжения на выходе которого определяется только порядком переключения вентилей инвертора, а форма тока зависит от характера нагрузки. Питание автономного инвертора осуществляется от источника напряжения;
Инвертор тока – это инвертор, на выходе которого форма тока определяется только порядком переключения вентилей инвертора, а форма напряжения зависит от характера нагрузки. Питание инвертора этого типа должно выполняться от источника тока;
Частотно-регулируемый привод (ЧРП) — система управления частотой вращения ротора асинхронного (или синхронного) электродвигателя. ЧРП состоит из электродвигателя и частотного преобразователя;
DC/AC Drive — это устройство, состоящее из выпрямителя, преобразующего переменный ток промышленной частоты в постоянный, и инвертора, преобразующего постоянный ток в переменный, требуемых напряжения и частоты;
DC/AC converter — устройство, преобразующие электрическую энергию источника напряжения постоянного тока в электрическую энергию переменного тока с изменением величины частоты и/или напряжения.
Устройство плавного пуска (УПП) – это электронное устройство, используемое для плавного пуска и торможения синхронных и асинхронных электродвигателей.
Низковольтное УПП — устройство плавного пуска работающее на низком напряжении до 660 В.
Высоковольтное УПП — устройство плавного пуска работающее на средних напряжениях до 15 кВ.
В различный устройство плавного пуска может называться также:
- Softstarter
- Софтстартер
- Устройство мягкого пуска
- Мягкий пуск
- Плавный пуск
- Тиристорный регулятор напряжения
Softstarter или Софтстартер — устройство плавного пуска;
Устройство мягкого пуска (мягкий пуск) – устройство плавного пуска;
Тиристорный регулятор напряжения – это устройство, предназначенное для регулирования частоты вращения электродвигателя и используется главным образом для управления пуском. Регулировка происходит в результате изменения напряжения, которое подведено к статору двигателя, и смены угла открывания тиристоров.
Регулировка происходит в результате изменения напряжения, которое подведено к статору двигателя, и смены угла открывания тиристоров.
Сетевой дроссель – устройство для сглаживания скачков напряжения в питающей сети и компенсации высших гармоник из сети в преобразователь и обратно.
Сглаживающий дроссель — устройство для компенсации высших гармоник из сети в преобразователь и обратно.
Радиочастотные фильтры (фильтры ЕМС) – устройство для снижения уровня помех в широком диапазоне частот 150 кГц-30мГц.
Дроссель du/dt или Фильтр du/dt — устройство компенсации импульсов напряжения на выходе преобразователя частоты. Применяется для защиты изоляции обмоток электродвигателя.
Моторный дроссель — устройство компенсация скачков напряжения на клеммах двигателя. В отличие от фильтров du/dt, моторный дроссель обладает существенно большей индуктивностью и применяется при больших длинах кабеля и большой частоте переключений.
Синусный фильтр — устройство для обеспечения синусоидального напряжения между клеммами электродвигателя. Обеспечивает комплексную защиту двигателя и частотного преобразователя.
Тормозной резистор – устройство обеспечения быстрого торможения, применяемое в составе привода с большой инерцией.
ШИМ, Широтно-импульсная модуляция — это способ кодирования аналогового сигнала путём изменения ширины (длительности) прямоугольных импульсов несущей частоты. Принцип ШИМ применяется в частотных преобразователях для получения питающего напряжения нужной частоты.
Устройства плавного пуска — что это и для чего? » Вcероссийский отраслевой интернет-журнал «Строительство.RU»
Итак, УПП — это механическое или электронное устройство (также существуют электромеханические модели), которое обеспечивает плавный разгон и торможение электродвигателя за счет параметрирования величины напряжения.
В момент пуска двигателя крутящий момент за считанные секунды может достигать 150-200%, а пусковой ток — 600-800% от номинального. Из-за этого в местной электросети может возникнуть просадка напряжения, что повлечет за собой аварийный останов двигателя, или, наоборот, резкое увеличение нагрузки на него. Устройство плавного пуска призвано предотвратить все эти негативные последствия. Благодаря плавному разгону привода УПП предотвращает соударения шестеренок внутри редукторов, проскальзывание клиновидных ремней, гидравлические удары в трубопроводах, колебания в конвейерных системах и так далее.
Из-за этого в местной электросети может возникнуть просадка напряжения, что повлечет за собой аварийный останов двигателя, или, наоборот, резкое увеличение нагрузки на него. Устройство плавного пуска призвано предотвратить все эти негативные последствия. Благодаря плавному разгону привода УПП предотвращает соударения шестеренок внутри редукторов, проскальзывание клиновидных ремней, гидравлические удары в трубопроводах, колебания в конвейерных системах и так далее.
На рынке существует множество производителей устройств плавного пуска, но продешевить и выбрать сомнительный прибор — значит выкинуть деньги на ветер. К выбору оборудования нужно подходить, исходя из конкретных задач. Например, корпорация Siemens предлагает дорогое, но весьма надежное оборудование, которое отличается многофункциональностью и широким модельным рядом. Вот основные серии подобных УПП: http://tehprivod.ru/katalog/ustroystva-plavnogo-puska. Устройства плавного пуска немецкого производства обеспечивают полную защиту электродвигателя от всех видов негативных воздействий нестабильных электросетей.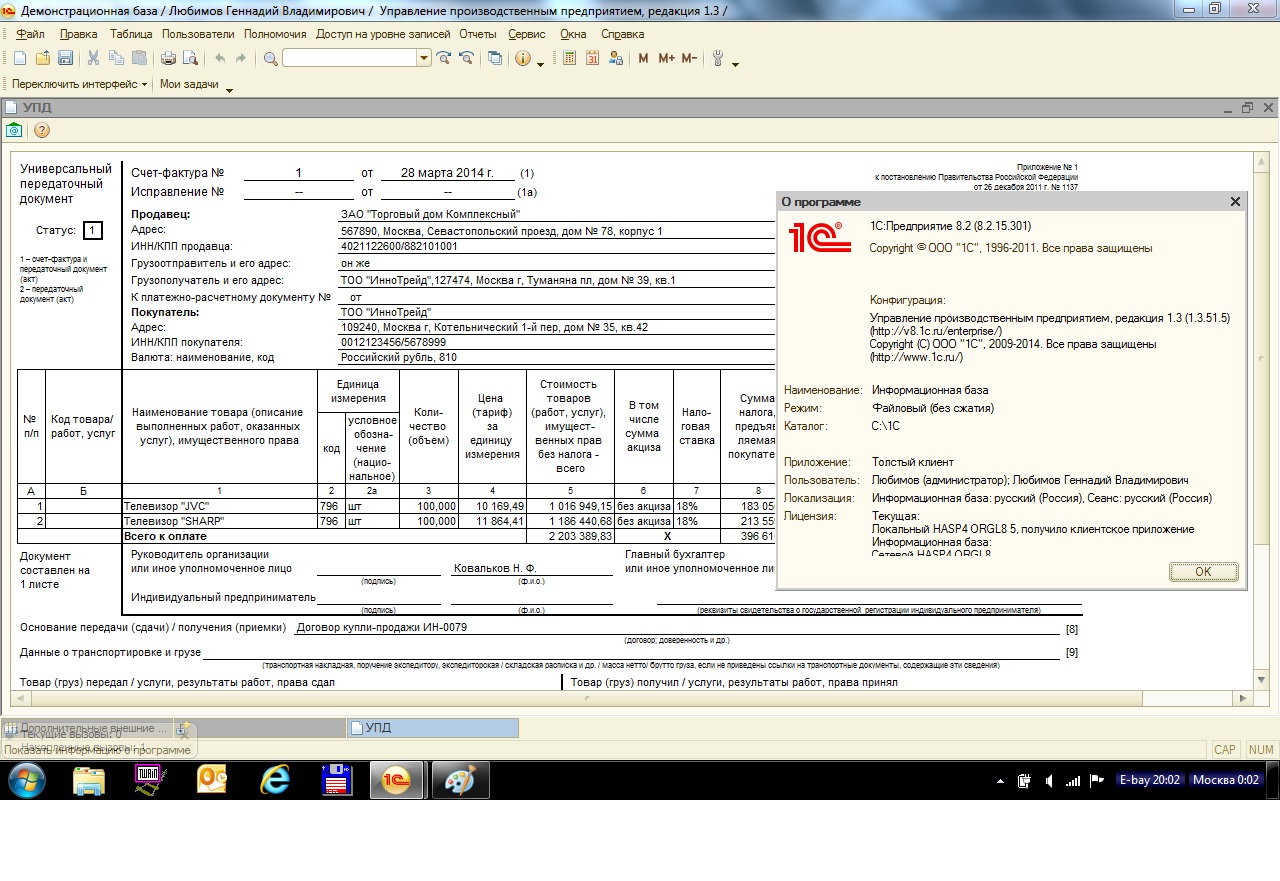 Недостатков у такого оборудования практически нет, а потому пускатели Siemens необходимо ставить только на самые ответственные участки производства.
Недостатков у такого оборудования практически нет, а потому пускатели Siemens необходимо ставить только на самые ответственные участки производства.
Производитель Prostar предлагает отличное соотношение цены и качества, но с меньшей универсальностью, чем оборудование Siemens. Для устройств плавного пуска тайваньской компании также характерна надежность, широкий модельный ряд и простое подключение, но с меньшими затратами, а значит такие пускатели целесообразно ставить на агрегаты среднего звена. Для менее ответственных участков с небольшими мощностями можно подобрать оборудование менее именитых китайских производителей. Как правило, их УПП дешевы, универсальны, но менее надежны. В случае поломки такое устройство можно быстро заменить запасным — низкая стоимость пускателя позволит сделать это без большого ущерба для бюджета.
Однако не будем сгущать краски, так как устройства плавного пуска, кто бы их не производил, действительно уберегают оборудование. Даже прибор среднего качества может предотвратить выход из строя дорогостоящего электродвигателя.
Виды запасов: что это, и что делать, чтобы все было хорошо
25 Ноября 2019
Учет товаров по видам запасов, особенности его реализации в программах 1C:Управление торговлей 11, Комплексная автоматизация 2 и 1C:ERP, а также отличия от «традиционного» партионного учета.
В конфигурациях предыдущего поколения, таких как Управление торговлей 10.3, Управление производственным предприятием 1.3 (В 1С:УПП при работе в режиме «Партионного учета») все необходимые данные по учету товаров (где лежат, кому принадлежат, в каком состоянии и т.д.) хранились в отдельных измерениях регистров, при этом движения регистров переформировывались каждый раз при перепроведении документов (рисунок 1).
Рисунок 1
Такой подход был удобен на небольших объемах данных, на которых было легко исправить возможные ошибки в учете, заменить некорректные данные корректными и т.д. Однако, на больших объемах данных, когда количество документов в месяц составляет несколько тысяч и более, использование стандартного механизма перепроведения, то есть восстановления последовательности документов, уже весьма проблематично и требовало большого количества времени на свою работу в монопольном режиме (иначе не избежать блокировок работы других пользователей).
Кроме того, этот подход требовал больших временных затрат при расчете встречного выпуска при производстве (из-за итерационного подхода) и сдерживал возможности указания большего количества аналитики в регистре, что требовало большего количества регистров для хранения этих данных.
Поэтому в новых решениях, таких как 1С:Управление торговлей 11( 11.0.х, 11.1.х), 1С:Комплексная автоматизация 1.1 и 1С:УПП 1.3 Фирма 1С приняла решение:
- расчет себестоимости списания разместить в процедуре закрытия месяца,
- начать использовать индексные справочники, которые объединили большой набор измерений регистра в одно измерение – ссылку на индексный справочник. Благодаря этому получалось значительное уменьшение размеров таблиц движений и итогов по регистрам учета себестоимости. Это была «Расширенная аналитика учета затрат» (рисунок 2).
Рисунок 2
Как видно на рисунке, в четырех реальных измерениях регистра хранится более 25 «ссылочных» измерений.
Дальнейшим развитием этого механизма послужил механизм учета ТМЦ «По видам запасов». Он в течении времени также менялся, поэтому рассмотрим современную версию.
Подход РАУЗ был в некотором смысле слишком глобальным (учет ТМЦ и затрат на одном большом регистре), и в современных решениях количество полей в индексных справочниках было заметно уменьшено, а главное – они были более логично сгруппированы (рисунок 3).
Рисунок 3
В «Аналитике учета номенклатуры» сгруппированы «управленческие» характеристики, а в «Виде запасов» — относящиеся к регламентированному учету: юридическое лицо, тип запасов, вид налогообложения и важная характеристика — «группа фин. учета», в которой и задается способ отражения по счетам учета (рисунок 4).
Рисунок 4
Вроде всё понятно и логично, но есть один нюанс. Для роста производительности, уменьшения случайных изменений учета при перепроведениях, а также для возможности ручного выбора – виды запасов в документах расхода теперь хранятся в отдельной скрытой табличной части, которая не перезаполняется при перепроведении документа (даже если она сформирована автоматически). Например, если поменять группу фин. учета в товаре и перепровести документы с его участием, это не повлечет изменений в учете, так как документы уже «запомнили» старый вариант (рисунок 5).
«Да здравствуют отрицательные остатки!».
Рисунок 5
Конечно, в 1С:Комплексная Автоматизация,2.4, 1С:ERP и 1C:Управление Торговлей, 11 есть инструмент, который может помочь исправить эту ситуацию. Он находится в закрытии месяца и выводится в том случае, если система считает, что он может помочь (рисунки 6 и 7).
Рисунок 6
Рисунок 7
Однако он может справиться не всегда. Например, не может решить проблему некорректных видов запасов в документах поступления. В этом случае приходится применять один из трех подходов:
- Править виды запасов вручную. Очень трудоемкий подход.
- Отменять проведение документов с очисткой видов запасов в документах поступления и потом их заново проводить Тут могут возникнуть сложности, если в базе есть контроль отрицательных остатков организаций или на складах.
 Кроме того, при групповом проведении документов в КА 4 и ЕРП документы часто блокируют сами себя из-за реализации части проведения в виде фоновых заданий.
Кроме того, при групповом проведении документов в КА 4 и ЕРП документы часто блокируют сами себя из-за реализации части проведения в виде фоновых заданий. - Очистить виды запасов обработкой у проведенных документов и потом перепровести их. Может потребоваться несколько итераций.
Мы рекомендуем третий подход, как самый быстрый из возможных.
Также в интернете встречаются расширения, включающие перезаполнение видов запасов при каждом перепроведении документов.
Статью подготовил Василий, старший специалист по внедрению 1С:Предприятие компании «ИнфоСофт»
Оригинал статьи на портале ИнфоСтарт
azeam / powerupp: простой графический интерфейс для UPP
Простой графический интерфейс для UPP (https://github.com/sibradzic/upp)
Это инструмент для простой настройки графического интерфейса (выбранного количества) значений в таблице AMD PowerPlay в Linux.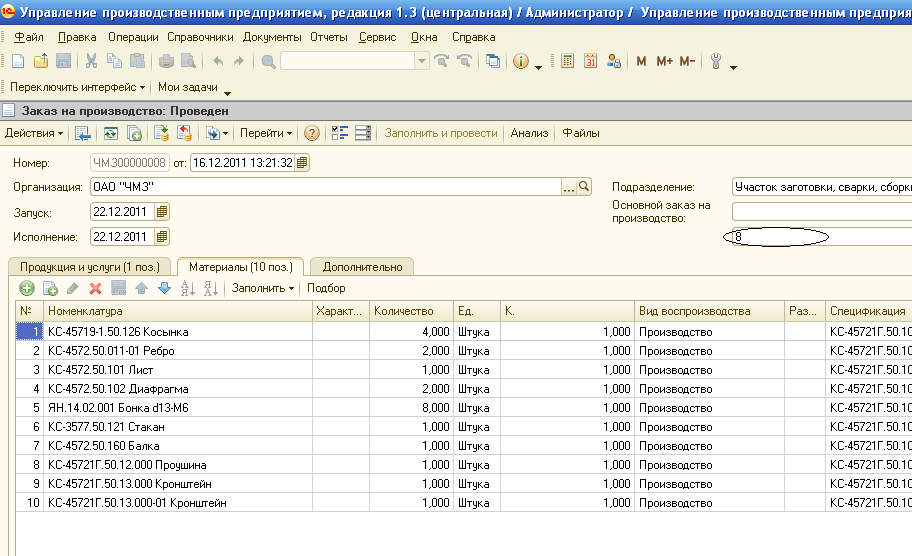 Графические карты AMD Navi 10 (серия Radeon 5000) и Big Navi (серия Radeon 6000) поддерживаются, но прочтите примечания ниже.
Графические карты AMD Navi 10 (серия Radeon 5000) и Big Navi (серия Radeon 6000) поддерживаются, но прочтите примечания ниже.
Причина создания этого заключается в том, что настройки OverDrive все еще не работали для карт Navi 10, когда я начал над ним работать (ядро <5.5). Благодаря sibradzic и его работе с UPP, можно в любом случае настроить параметры (и нет необходимости устанавливать параметры ядра ppfeaturemask ), но может быть трудно интерпретировать огромное количество параметров из файла pp_table , поэтому это GUI.
Для тех из вас, кто владеет картой Navi и использует ядро <5.5 и хочет разогнать / понизить напряжение и т. Д., Или если вы хотите повозиться со значениями, недоступными в настройках OverDrive для более экстремального или точного разгона - это приложение может облегчить процесс.
Несмотря на простой внешний вид, это мощный инструмент, который потенциально может повредить вашу видеокарту, если не принять меры. Он находится на ранней стадии разработки, и это моя первая попытка кодирования на C (вероятно, множество примеров плохого кода, но со временем я постараюсь улучшить базу кода). В моей системе он полностью функционален в своем текущем состоянии, но используйте его на свой страх и риск!
В моей системе он полностью функционален в своем текущем состоянии, но используйте его на свой страх и риск!
Если вы считаете, что некоторые важные настройки отсутствуют / должны быть реализованы (для управления вентилятором проверьте CoreCtrl или radeon-profile, не беспокойтесь об этом) или у вас есть какие-либо другие проблемы, сообщите о проблеме.Лично у меня мало опыта разгона, но я думаю, что включенные настройки должны быть самыми важными. Для оптимальных настроек вам нужно будет поискать в другом месте или поэкспериментировать самостоятельно, я использую его только для снижения напряжения моего 5700 XT для снижения шума и энергопотребления.
Примечания
Начиная с ядра Linux 5.10 кажется необходимым установить флаг ppfeaturemask , чтобы настроить ограничение мощности, добавьте строку amdgpu.ppfeaturemask = 0xffffffff в качестве флага загрузки ( sudo nano / etc / default / grub при использовании GRUB), обновите sudo update-grub и перезагрузитесь. Для старых ядер вам может потребоваться более старая прошивка, см. №3.
Для старых ядер вам может потребоваться более старая прошивка, см. №3.
У карты AMD 5600 XT есть некоторые строгие ограничения прошивки, которые, кажется, не позволяют установить часы Gfx выше стандартных настроек с помощью pp_table . Однако можно настроить другие параметры, а затем разогнать до предела OverDrive с помощью других инструментов (OverDrive). Более подробное обсуждение этого вопроса см. В выпуске №1.
В настоящее время, похоже, существуют определенные ограничения для карт серии 6000, см. # 21 для получения дополнительной информации.
Зависимости
GTK3 (dev)
GCC
Python 3.6+
pip
UPP 0.0.7+ (0.0.8+ для Big Navi), устанавливается как пакет pip: pip3 install upp
Для Ubuntu:
-
sudo apt-get install libgtk-3-dev build-essential python3 python3-pip -
pip3 установить upp
Установка
git clone https://github.  com/azeam/powerupp.git
com/azeam/powerupp.git
cd powerupp
make
sudo make install
Удалить
sudo make uninstall
Использование
Команда install установит файл.файл запуска рабочего стола, который должен быть загружен Gnome и т. д. Если у вас возникли проблемы и вам нужен более подробный вывод, вы также можете запустить приложение с терминала ( powerupp ).
PowerUPP поддерживает несколько графических карт, выберите карту, которую вы хотите настроить, в раскрывающемся меню. «Загрузить активные» настройки при первом использовании приложения, это прочитает и переведет содержимое вашего файла (ов) pp_table . Эти значения будут сохранены как «значения по умолчанию» в папке / home / USER /.config / powerupp / и позже может быть использован для сброса значений на stock. Рекомендую не редактировать этот файл вручную.
Если вы установили собственные значения powerplay перед использованием этой программы или удалили файл значений по умолчанию после его создания, будьте осторожны, «безопасные» пределы по умолчанию могут быть неправильными. «Безопасные» пределы устанавливаются в соответствии со значениями, представленными в таблице AMD: s OverDrive (такой же, как в Wattman и т. Д.), Если они доступны.
«Безопасные» пределы устанавливаются в соответствии со значениями, представленными в таблице AMD: s OverDrive (такой же, как в Wattman и т. Д.), Если они доступны.
Когда вы примените свои настройки, они сразу же вступят в силу, но не сохранят перезагрузку.Обратите внимание, что все отображаемые значения будут записаны в pp_table при нажатии «Apply current», рекомендуется изменять одно значение за раз.
Чтобы сохранить текущие настройки активными после перезагрузки, используйте параметр «Постоянное сохранение». Это создаст сценарий bash и файл правил udev, который загружает сценарий при запуске системы. Обратите внимание, что «Постоянное сохранение» будет считывать и сохранять значения, отображаемые в приложении, а не текущие активные настройки. Удаление приложения с помощью команды sudo make uninstall из каталога git также отключит и удалит сценарии запуска, если они включены, но не сохраненные профили или значения по умолчанию.
Партнерская программа WEF Utility
Есть дополнительная плата за создание UPP?
Нет, за создание UPP плата не взимается. Каждый член является индивидуальным членом — руководителем, профессионалом, оператором и молодым специалистом.
Есть ли ограничение на количество членов, которое вы можете иметь в UPP?
- Коммунальные предприятия должны иметь как минимум пять сотрудников для участия в программе.
- Нет ограничений на количество членов WEF, которые могут быть добавлены к вашей учетной записи UPP.
Что, если кто-то покинет коммунальное предприятие в середине цикла членства?
Коммунальная компания оплатила членство. Следовательно, если утилита уведомит нас, WEF прекратит членство. Затем WEF переведет оставшиеся членские взносы на счет другого сотрудника.
Что произойдет с индивидуальным членством, если сотрудник покинет назначенное коммунальное предприятие?
Когда сотрудники WEF узнают, что кто-то покинул указанное коммунальное предприятие, делается все возможное, чтобы связаться с этим членом и уведомить его о том, что его членство больше не оплачивается UPP. Важно, чтобы участники зашли в свой профиль членства в WEF и указали личные адреса электронной почты и домашние адреса. Это позволит сотрудникам WEF уведомлять его / ее о любых изменениях.
Важно, чтобы участники зашли в свой профиль членства в WEF и указали личные адреса электронной почты и домашние адреса. Это позволит сотрудникам WEF уведомлять его / ее о любых изменениях.
Должны ли вы индивидуальные члены вступать в членскую ассоциацию?
Да, все члены WEF в UPP должны принадлежать к членской ассоциации.
Все ли ассоциации-члены участвуют в программе партнерства с коммунальными предприятиями?
В настоящее время пять ассоциаций-членов не принимают программы UPP.
- Водная ассоциация штата Аризона
- Калифорнийская ассоциация водной среды
- Ассоциация водной среды Нью-Джерси
- Quebec –Réseau Environment
- Ассоциация водной среды Южной Каролины
Что делать, если цикл членства сотрудников отличается от членского цикла UPP?
Персонал ВЭФ будет рассматривать каждый индивидуальный цикл членства. Они будут пропорционально оценивать индивидуальный цикл членства, чтобы он соответствовал циклу UPP.

Как происходит продление UPP?
Каждого UPP просят назначить контактное лицо UPP. За два месяца до продления WEF отправит контактному лицу UPP счет. Эти временные рамки должны дать контактному лицу достаточно времени, чтобы просмотреть счет и внести какие-либо изменения. Если есть изменения, Контактное лицо отправляет изменения обратно в WEF, и пересмотренный счет создается и отправляется обратно Контактному лицу.
Получит ли отдельный член уведомление о продлении?
Индивидуальный участник не должен получать уведомление о продлении.Одним из преимуществ наличия UPP является то, что многочисленные сотрудники не будут подавать заявления о продлении членства в разное время в течение года.
Получит ли индивидуальный член членскую карту WEF?
Да, каждый член WEF является индивидуальным членом, и членская карта будет отправлена после оплаты счета UPP.
PCSAO — Программа партнерства университетов
Программа партнерства с университетами (UPP) — это уникальное сотрудничество между Департаментом занятости и семейных услуг штата Огайо, восемью государственными школами социальной работы штата Огайо, PCSAO, государственными агентствами по обслуживанию детей штата Огайо (PCSA) и Программой обучения благополучию детей штата Огайо.
PCSAO не управляет этой программой, но активно участвует в ее поддержке. Для получения дополнительной информации свяжитесь с Трейси Притчард.
Целью UPP является усиление профессиональных кадров в области защиты детей в Огайо за счет увеличения числа хорошо подготовленных выпускников со степенью бакалавра и магистра в области социальной работы, которые поступают в эту сферу. Исследования показывают, что те, кто заканчивает такую программу, как UPP, остаются дольше и более довольны, чем те, кто не завершил такую программу до трудоустройства.
студентов UPP проходят специальные курсы по защите детей, проходят стажировку в PCSA и по окончании получают частичное возмещение стоимости обучения после принятия на работу в PCSA штата Огайо. Тем, кто завершает программу, возможно, не придется проходить необходимое базовое обучение (102 часа), которое все другие новые соцработники должны пройти в течение первого года работы.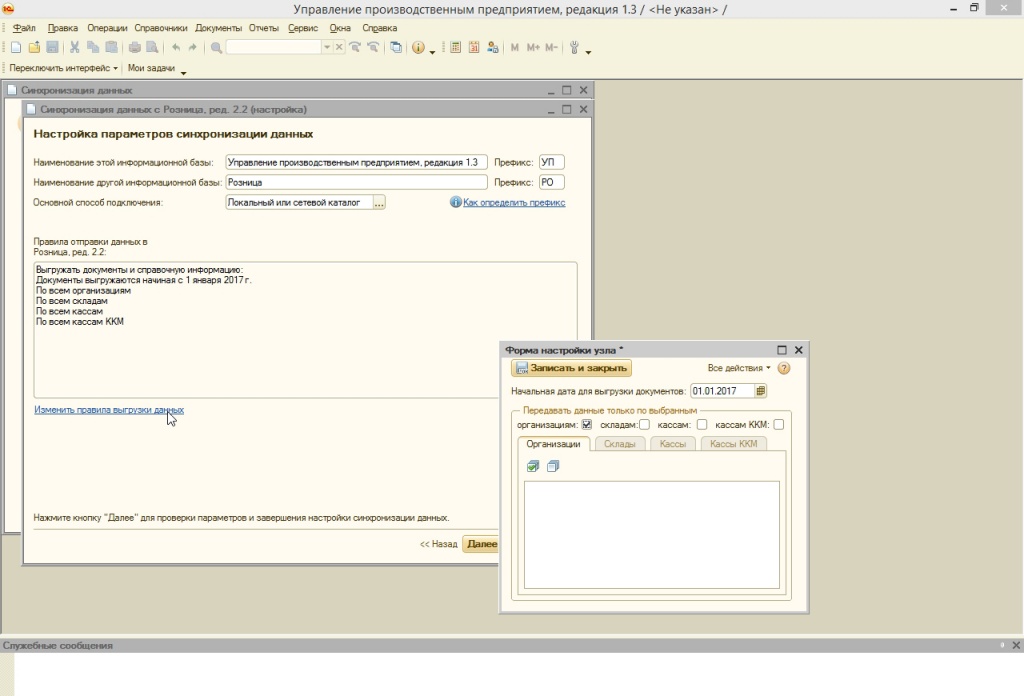 Выпускники UPP стали регулярно искать вакансии социальных работников PCSA.
Выпускники UPP стали регулярно искать вакансии социальных работников PCSA.
Для абитуриентов
Для студентов и выпускников
Следующие ресурсы были разработаны Комитетом по кадрам и безопасности PCSAO для помощи выпускникам UPP в сравнении требований к работе и льгот в различных агентствах.Опыт показывает, что одно агентство может предлагать больше по заработной плате, но предлагать меньше в своем общем пакете из-за разных взносов на дополнительные пособия и различных требований и возмещения расходов в таких обстоятельствах, как «дежурство по вызову». Студенты могут использовать «Вопросы сравнения преимуществ агентств», чтобы узнать, что им следует учесть, прежде чем заполнять «Рабочий лист оценки предложений».
Для агентств
Оценка программы
рабочих групп | Деловые партнерства и услуги университетов
Состоящие из заинтересованных сторон сообщества университетского городка, рабочие группы по конкретным категориям формируются для разработки запросов на предложения (RFP) для каждой области партнерства и для рекомендации перспективных партнерств Консультативному комитету. UPP объединяет уникальную группу для изучения каждой категории, привлечения экспертных знаний в кампусе и предоставления членам сообщества университетского городка возможности высказать свое мнение о предполагаемых категориях и партнерствах. Заинтересованные стороны могут включать преподавателей, сотрудников, студентов, университетские подразделения, выпускников и других.
UPP объединяет уникальную группу для изучения каждой категории, привлечения экспертных знаний в кампусе и предоставления членам сообщества университетского городка возможности высказать свое мнение о предполагаемых категориях и партнерствах. Заинтересованные стороны могут включать преподавателей, сотрудников, студентов, университетские подразделения, выпускников и других.
Члены Рабочей группы по напиткам
Студенческий союз ASUC: Bahar Navab
Cal Питание: Кристофер Хеннинг
Представитель факультета: Кристин Мэдсен
Представитель аспиранта: Мартин Сирон
Межуниверситетская легкая атлетика: Джо Малфорд
Управление устойчивого развития: Кира Столл
Отдел по делам студентов Развитие бизнеса: Кирк Роблес
Цепочка поставок Руководство: Стейси Темплман
Представитель студентов бакалавриата: Селена Мелгоза
Медицинские услуги университета: Ким Гесс
Члены страховой рабочей группы
Ассоциация выпускников Калифорнийского университета: Мэтт Тервиллигер
Представитель факультета: Нэнси Уоллес
Представитель аспирантов: Аф Эрнандес
Лирфилд / Легкая атлетика: Энди МакДауэлл
Люди и культура: Брианна Вандре
Дела со студентами: Джо Уотц
Управление цепочками поставок: Стейси Темплман
Студент бакалавриата Представитель: Маргарет МакГилл
Искусство и дизайн: Массимо Пакчионе
Ассоциация выпускников Калифорнийского университета: Мэтт Тервиллигер
Представители факультета: Янгос Папанастасиу
Представитель аспирантов: Джейми Линкофф *; замена TBA
Отдел кадров: Джеймс Дудек
Лирфилд: Энди МакДауэлл
Парковка и транспорт: Шеймус Уилмот
Дела со студентами: Джо Уотц
Управление цепочкой поставок: Стейси Темплман
Технология: Билл Эллисон
Представитель студентов бакалавриата: Анна Уитни
Члены рабочей группы по одежде
Студенческий союз ASUC: Дэрил Ансель
Деловые контракты и защита бренда: Мишель Хафф
Ассоциация выпускников Калифорнийского университета: Мэтт Тервиллигер
Связь и связи с общественностью: Рам Капур
Представитель факультета: Келли МакЭлхани
Представитель аспирантов: Лизз Нимейер * и Джейми Линкофф *; замена TBA
Межвузовская легкая атлетика: Ашвин Пури *; Джастин Панарезе *; Джо Мулфорд
Спортивный отдых: Джо Уотц
Недвижимость: Хелен Левай
Skydeck: Кэролайн Виннетт
Представитель студентов бакалавриата: Радж Бхаргава
Развитие университетов и отношения с выпускниками: Сильвия Берхуис
Члены рабочей группы по кофе
Студенческий союз ASUC: Сьюзи Халпин * и Дэрил Ансел
Ассоциация выпускников Калифорнийского университета: Мэтт Тервиллигер
Представитель факультета: Кейт Гиллесс
Представитель аспирантов: Марк Шаперо
Межуниверситетская легкая атлетика: Джастин Панарезе *; Джо Малфорд,
Управление устойчивого развития и энергетики: Кира Столл
Спортивный отдых: Джо Уотц
Недвижимость: Хелен Левай
Управление цепочкой поставок: Стейси Темплман
Представитель студентов бакалавриата: Джиана Амадор * и Уэс Адриансон *, в настоящее время TBD
Медицинские услуги университета: Триш Ратто * и Кори Эванс
Члены банковской рабочей группы
Студенческий союз ASUC: Келси Финн *, Тереза Аджари * и Дэрил Ансел
Легкая атлетика: Ашвин Пури *; Джастин Панарезе *; Джо Малфорд
Cal 1 Card: Мэри-Энн Спенсер Коган
Калифорнийская ассоциация выпускников: Мэтт Тервиллигер
Преподаватель: Нэнси Уоллес
Представитель аспирантов: Энтони Абрил * и Джейми Линкофф
Поставки: Стейси Темплман
Спортивный отдых: Джо Уотц
Представитель студентов бакалавриата : Радж Бхаргава
Члены рабочей группы Brita
Промышленная категория больше не активна.
Студенческий союз ASUC: Сюзи Халпин * и Дэрил Ансель
Ассоциация выпускников Калифорнийского университета: Мэтт Тервиллигер
Представитель аспирантов: Марк Шаперо
Межуниверситетская легкая атлетика: Джастин Панарезе
Управление устойчивого развития и энергетики: Кира Столл
Недвижимость: Брайан Мэйн *
Развлекательные виды спорта: Джо Уотц
Представитель студентов бакалавриата: Крис Ямас
Медицинские услуги университета: Триш Ратто * и Кори Эванс
Члены Рабочей группы по коммуникациям
Промышленная категория больше не активна.Студенческий союз ASUC: Дарли Ансел
Искусство и дизайн: Сьюзан Миллер
Ассоциация выпускников Калифорнийского университета: Мэтт Тервиллигер
Инженерный колледж: Энтони Сент-Джордж
Декан по делам студентов: Дэвид Сурратт
Образовательные услуги: Дженн Стрингер
Факультет Представитель: Джеффри Бокор,
, аспирант Представитель: Мэтью Хан,
Информационная служба и технологии: Лайл Невелс,
Межуниверситетская легкая атлетика: Эшвин Пури * и Джастин Панарезе
Недвижимость: Хелен Левай
Спортивный отдых: Джо Уотц
Управление цепочкой поставок: Стейси Темплман
Technology : Уильям Эллисон
Телекоммуникации: Ховард Ревел и Майкл Грин
Дополнительный номер Калифорнийского университета в Беркли: Роберт Дэвид
Представитель студентов бакалавриата: Эмбер Норори
Члены рабочей группы авиакомпаний
Промышленная категория больше не активна.
Искусство + дизайн: Ральф Скала
Ассоциация выпускников Калифорнийского университета: Мэтт Тервиллигер
Представитель факультета: Джеффри Пеннингтон
Представитель аспирантов: Дэвид Братт
Межвузовская легкая атлетика / Лирфилд: Джастин Панарезе / Джо Малфорд
По делам студентов: Кэрон Ламарш
Путешествие: Дэн Парнас
Представитель студентов бакалавриата: Monsoon Pabrai
Офис президента Калифорнийского университета: Дебра Алмейсон
Члены Рабочей группы по солнечной энергии
I Промышленная категория больше не действует.Berkeley Energy and Resources Collaborative:
Chris Reilly *Калифорнийская ассоциация выпускников: Мэтт Тервиллигер
Межуниверситетская легкая атлетика: Джастин Панарезе
Зал науки Лоуренса: Сьюзан Грегори
Управление устойчивого развития: Кира Столл
Управление цепочкой поставок: Рич Тейлор
* Участвует в процессе переговоров по контракту. Человек может не входить в текущую рабочую группу или больше не работать в университете.
Человек может не входить в текущую рабочую группу или больше не работать в университете.
Увеличение мощности — сок Upp
Супер заряженная формула, специально для тех, кто был истощен жизнью.
Power Upp может быть отличным способом сформировать возвращение и добавить энергии и сосредоточиться на тренировках, если вы какое-то время были неактивны.
На рынке есть много продуктов для повышения тестостерона — так какой из них выбрать? Мы вернулись к основам и сделали это простым, и наши эксперты выбрали стимуляторы тестостерона CORE, которые нам действительно нужны. Так что не нужно добавлять в эту формулу ничего необычного.
Эта формула содержит соединения, которые, как считается, увеличивают выработку тестостерона.
Основным ингредиентом этой формулы является ОЧЕНЬ МОЩНАЯ доза Horny Goat Weed (HGW), также известного как Epimedium sagittatum. Роговой козий сорняк уже давно используется в традиционной восточной медицине.
В целом известно, что HGW улучшает кровоток, ингибируя человеческий PDE5, вещество, которое блокирует эрекцию. Также известно, что улучшенные свойства HGW способствуют повышению уровня выработки тестостерона.
ВPower Upp также есть популярный Тонгкат Али. Известно, что этот экстракт увеличивает мышечную массу и силу, ускоряя потерю жира у здоровых людей, занимающихся спортом, и, таким образом, может считаться естественным эргогенным средством для спортсменов и людей, сидящих на диете
Мака, анаболическая трава, которая может сильно повысить выработку тестостерона и поддержать рост мышц, включена в эту формулу, чтобы помочь удалить радикалы в организме и повысить спортивные результаты за счет балансировки гормонов.
Порошок Polypodium vulgare, древнее лекарство, используемое на протяжении тысячелетий во всем мире, добавлен в эту смесь, чтобы обеспечить устойчивый баланс вашей иммунной системе и психическому здоровью.
Многообещающие исследования показывают, что Saw Palmetto может помочь повысить уровень тестостерона, улучшить здоровье простаты и уменьшить воспаление — так что щепотка этого вещества будет иметь большое значение для повышения эффективности Power Upp.
Муира Пуама стабильно завоевывает репутацию средства повышения либидо и улучшения гормональной коррекции. Некоторые клинические исследования показали, что экстракты муира пуамы полезны для центральной нервной системы и обладают адаптогенным и антистрессовым действием. Специально приготовленный экстракт корня муира пуама был запатентован за его способность «снимать физическую и умственную усталость».
Мы добавили в эту добавку чудесную траву Pansang Ginseng, которая, как известно, укрепляет вашу сердечно-сосудистую, иммунную и нервную системы. Похоже, что он увеличивает силу и выносливость за счет улучшения функции надпочечников во время тренировок. Он помогает организму адаптироваться к стрессам, как физическим, так и эмоциональным, и многочисленные исследования подтверждают его способности. Фактически, Всемирная организация здравоохранения заявляет, что клинические данные подтверждают использование женьшеня для улучшения умственных и физических возможностей в случаях слабости, истощения, утомления и потери концентрации.
Фактически, Всемирная организация здравоохранения заявляет, что клинические данные подтверждают использование женьшеня для улучшения умственных и физических возможностей в случаях слабости, истощения, утомления и потери концентрации.
L-аргинин, также включенный в правильную дозу для Power Upp, имеет два эффекта: он превращается в оксид азота и помогает организму вырабатывать белок. Эти эффекты дают L-аргинину ряд потенциальных преимуществ, которые варьируются от здоровья сердца и боли в груди до помощи в наращивании мышц, , заживлении ран и улучшении мужской фертильности.
Разработка и применение системы контрселективной замены генов на основе upp для изучения S-слоя белка SlpX Lactobacillus acidophilus NCFM
РЕФЕРАТ
Анализ генома in silico Lactobacillus acidophilus NCFM в сочетании с предполагаемыми исследованиями экспрессии генов гены и регуляторные сети, которые потенциально важны для выживания, устойчивости и активности этого организма в желудочно-кишечном тракте. Корреляция ключевых генотипов с фенотипами требует эффективной системы замены генов. В этом исследовании было изучено использование урацилфосфорибозилтрансферазы (UPRTase) upp NCFM L. acidophilus в качестве маркера контрселекции для положительного отбора рекомбинантов, выделившихся из хромосомной интеграции плазмид на основе pORI. Был сконструирован изогенный мутант, несущий делецию гена upp и устойчивый к 5-фторурацилу (5-FU), токсичному аналогу урацила, который также является субстратом для UPRTase.Контролируемый интеграционный вектор на основе pORI размером 3,0 т.п.н., несущий кассету экспрессии upp , pTRK935, был сконструирован и введен в хозяина Δ upp , несущего вспомогательную плазмиду pTRK669. Внехромосомная репликация pTRK935 дополняла мутировавший хромосомный аллель upp и восстанавливала чувствительность к 5-FU. Этот фон хозяина обеспечивает платформу для двухэтапной стратегии интеграции и удаления плазмиды, которая позволяет отобрать рекомбинанты без плазмид с диким типом или мутантным аллелем целевого гена в присутствии 5-FU.
Корреляция ключевых генотипов с фенотипами требует эффективной системы замены генов. В этом исследовании было изучено использование урацилфосфорибозилтрансферазы (UPRTase) upp NCFM L. acidophilus в качестве маркера контрселекции для положительного отбора рекомбинантов, выделившихся из хромосомной интеграции плазмид на основе pORI. Был сконструирован изогенный мутант, несущий делецию гена upp и устойчивый к 5-фторурацилу (5-FU), токсичному аналогу урацила, который также является субстратом для UPRTase.Контролируемый интеграционный вектор на основе pORI размером 3,0 т.п.н., несущий кассету экспрессии upp , pTRK935, был сконструирован и введен в хозяина Δ upp , несущего вспомогательную плазмиду pTRK669. Внехромосомная репликация pTRK935 дополняла мутировавший хромосомный аллель upp и восстанавливала чувствительность к 5-FU. Этот фон хозяина обеспечивает платформу для двухэтапной стратегии интеграции и удаления плазмиды, которая позволяет отобрать рекомбинанты без плазмид с диким типом или мутантным аллелем целевого гена в присутствии 5-FU. Эффективность системы была продемонстрирована делецией в рамке считывания гена slpX (LBA0512), кодирующего новый секретируемый белок 51 кДа, связанный с комплексом S-слоя L. acidophilus . Полученный мутант Δ slpX показал более низкие скорости роста, повышенную чувствительность к додецилсульфату натрия и большую устойчивость к желчи. В целом, эта улучшенная система замены генов представляет собой ценный инструмент для исследования механизмов, лежащих в основе пробиотической функциональности L.ацидофилус .
Эффективность системы была продемонстрирована делецией в рамке считывания гена slpX (LBA0512), кодирующего новый секретируемый белок 51 кДа, связанный с комплексом S-слоя L. acidophilus . Полученный мутант Δ slpX показал более низкие скорости роста, повышенную чувствительность к додецилсульфату натрия и большую устойчивость к желчи. В целом, эта улучшенная система замены генов представляет собой ценный инструмент для исследования механизмов, лежащих в основе пробиотической функциональности L.ацидофилус .
Lactobacillus acidophilus NCFM — это коммерчески известная пробиотическая бактерия, которая широко используется в пищевых добавках, в молоке и кисломолочных продуктах (47). Этот штамм, изначально являвшийся кишечным изолятом человека с 1970-х годов (8), с тех пор тщательно исследовался на предмет различных желаемых признаков. В связи с важностью понимания молекулярных механизмов, участвующих в функциях пробиотиков, была определена полная последовательность генома L. acidophilus NCFM (2).Последовательность генома NCFM служит планом для идентификации in silico локусов генов-кандидатов и генных регуляторных сетей, которые могут играть важную роль в выживании и взаимодействиях с хозяином этого микроорганизма в желудочно-кишечном тракте (ЖКТ), включая гены, участвующие в кислотной толерантности (5). ), толерантность к желчи (38, 42), факторы приверженности (20), восприятие окружающей среды и реакция (7), использование пребиотических сахаров (9), биосинтез полисахаридов, разложение оксалатов (6) и продукция бактериоцина (22).Кроме того, продолжающиеся исследования экспрессии генов на микрочипах выявили конкретные представляющие интерес наборы генов, которые исследуются дальше. Фундаментальный подход к установлению функциональных ролей важных характеристик генов включает инактивацию генов и последующий фенотипический анализ связанных изогенных мутантов. Следовательно, эффективная система нокаута генов является ценным генетическим инструментом для корреляции ключевых генотипов с фенотипами и для функциональной характеристики генов, связанных с пробиотическими атрибутами L.
acidophilus NCFM (2).Последовательность генома NCFM служит планом для идентификации in silico локусов генов-кандидатов и генных регуляторных сетей, которые могут играть важную роль в выживании и взаимодействиях с хозяином этого микроорганизма в желудочно-кишечном тракте (ЖКТ), включая гены, участвующие в кислотной толерантности (5). ), толерантность к желчи (38, 42), факторы приверженности (20), восприятие окружающей среды и реакция (7), использование пребиотических сахаров (9), биосинтез полисахаридов, разложение оксалатов (6) и продукция бактериоцина (22).Кроме того, продолжающиеся исследования экспрессии генов на микрочипах выявили конкретные представляющие интерес наборы генов, которые исследуются дальше. Фундаментальный подход к установлению функциональных ролей важных характеристик генов включает инактивацию генов и последующий фенотипический анализ связанных изогенных мутантов. Следовательно, эффективная система нокаута генов является ценным генетическим инструментом для корреляции ключевых генотипов с фенотипами и для функциональной характеристики генов, связанных с пробиотическими атрибутами L.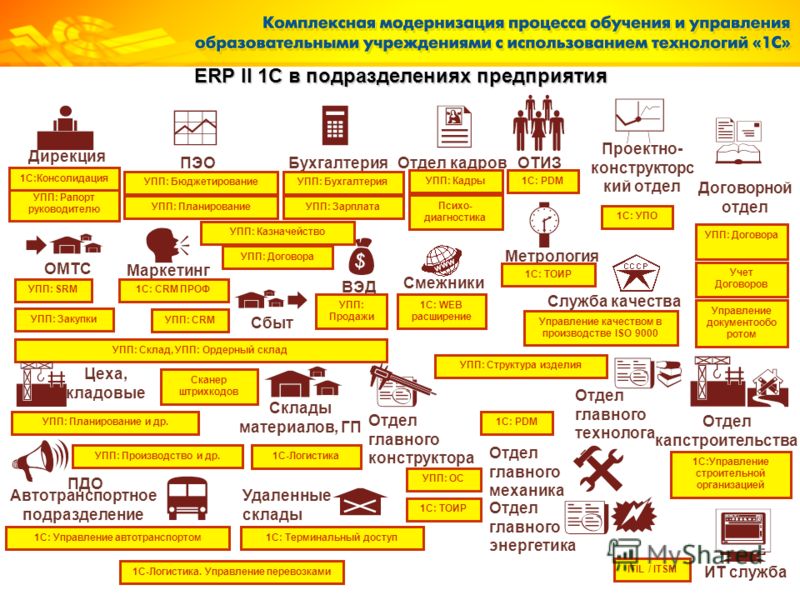 ацидофилус .
ацидофилус .
Разработка системы сайт-направленной лактококковой хромосомной интеграции Лоу и соавторами (34), включающая одновременное использование нерепликативного вектора, полученного из pWV01, с широким спектром хозяев (Ori + RepA —) или так называемого Вектор на основе pORI и термочувствительная вспомогательная плазмида pVE6007 (35), которая обеспечивает repA в trans для условной репликации плазмид на основе pORI, значительно облегчили генетические исследования различных грамположительных бактерий.Эта стратегия нокаута гена была адаптирована для использования в L. acidophilus и L. gasseri с альтернативной вспомогательной плазмидой, pTRK669, которая обеспечивает более высокий допустимый температурный диапазон для термофильных лактобацилл (46). Эта система с тех пор успешно использовалась для создания множества производных хромосомных вставок (1, 5, 9, 20, 23, 26, 38, 53). Тем не менее, стабильность инсерционных мутаций после гомологичной рекомбинации с одним кроссовером требует поддержания селекции антибиотиков, и один и тот же маркер селекции нельзя использовать для введения множественных мутаций в штамм. Более того, инсерционная инактивация специфической мишени в опероне может иметь полярные эффекты на нижележащие области. Эти ограничения были преодолены путем конструирования безмаркерных делеций генов посредством процесса гомологичной рекомбинации с двойным кроссовером, включающего интеграцию и вырезание плазмиды, где аллель дикого типа заменяется мутантным аллелем, несущим внутреннюю делецию в гене-мишени (6, 42). Однако из-за низкой эффективности удаления плазмиды и аллельной замены и отсутствия селектируемого маркера для обнаружения этих событий часто требуется обширный скрининг для выделения желаемых рекомбинантов.
Более того, инсерционная инактивация специфической мишени в опероне может иметь полярные эффекты на нижележащие области. Эти ограничения были преодолены путем конструирования безмаркерных делеций генов посредством процесса гомологичной рекомбинации с двойным кроссовером, включающего интеграцию и вырезание плазмиды, где аллель дикого типа заменяется мутантным аллелем, несущим внутреннюю делецию в гене-мишени (6, 42). Однако из-за низкой эффективности удаления плазмиды и аллельной замены и отсутствия селектируемого маркера для обнаружения этих событий часто требуется обширный скрининг для выделения желаемых рекомбинантов.
Одним из практических подходов к решению этой проблемы является включение контрселективных генетических маркеров в системы аллельного замещения для облегчения выделения производных, свободных от плазмид, после второго события рекомбинации. Среди установленных контрселективных маркеров — ген Bacillus subtilis sacB (кодирующий левансахаразу), который приводит к чувствительности к сахарозе (44), и ген galK (кодирующий галактокиназу), который опосредует токсичность галактозы или аналога галактозы (39, 49).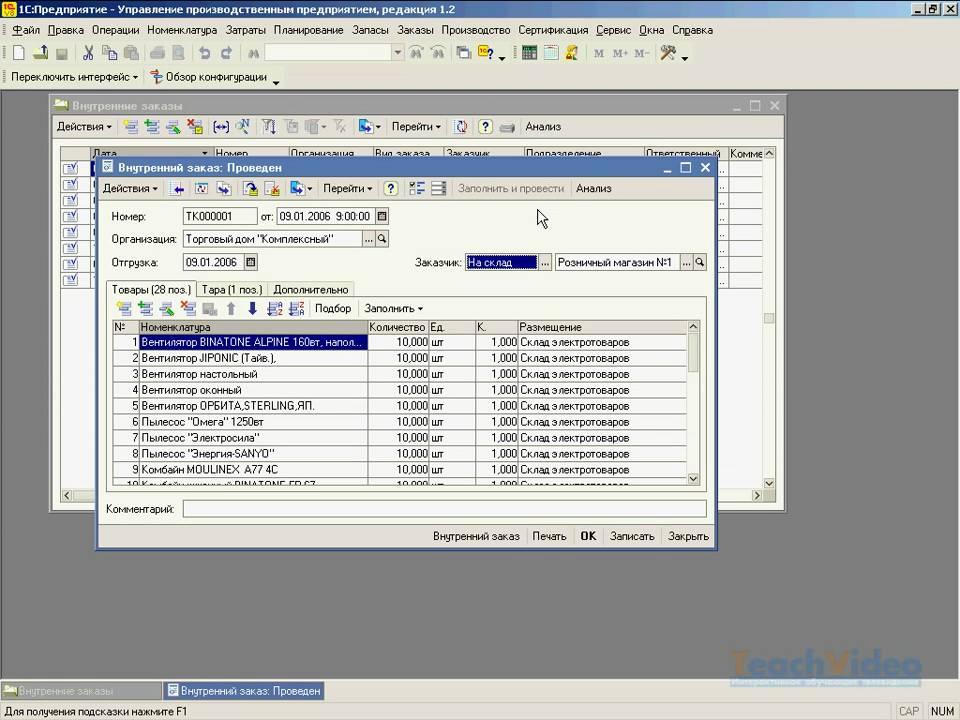 ) у меродиплоидов, не претерпевших второго гомологичного кроссовера.Гены, участвующие в путях спасения пуринов и пиримидинов, в частности гены, кодирующие фосфорибозилтрансферазы (PRTases), такие как upp (кодирует урацил PRTase [UPRTase]), hprT (кодирует гипоксантин PRTase), pyrE2 PRTase) и pryF / ura3 (кодирующий оротидин-5′-фосфатдекарбоксилазу) также являются контрселективными маркерами, которые широко используются в системах нокаута бактериальных, архейных и эукариотических генов (14, 15, 24, 25, 33, 41 , 43, 54).Эти PRTазы превращают предварительно сформированные пуриновые и пиримидиновые основания в соответствующие монофосфаты нуклеотидов для биосинтеза нуклеотидов de novo. Присутствие аналога основания, которое также распознается как субстрат целевой PRTase, может быть летальным для клеток, когда основание включено в предшественники нуклеотидов, тогда как на мутант с нефункциональной PRTase меньше влияет токсичность аналога основания. Подход встречного отбора включал использование вектора интеграции, который обеспечивает эктопическую экспрессию гена, кодирующего PRTase, в фоновом хозяине, дефектном по PRTase, для нацеленных на ген делеций.
) у меродиплоидов, не претерпевших второго гомологичного кроссовера.Гены, участвующие в путях спасения пуринов и пиримидинов, в частности гены, кодирующие фосфорибозилтрансферазы (PRTases), такие как upp (кодирует урацил PRTase [UPRTase]), hprT (кодирует гипоксантин PRTase), pyrE2 PRTase) и pryF / ura3 (кодирующий оротидин-5′-фосфатдекарбоксилазу) также являются контрселективными маркерами, которые широко используются в системах нокаута бактериальных, архейных и эукариотических генов (14, 15, 24, 25, 33, 41 , 43, 54).Эти PRTазы превращают предварительно сформированные пуриновые и пиримидиновые основания в соответствующие монофосфаты нуклеотидов для биосинтеза нуклеотидов de novo. Присутствие аналога основания, которое также распознается как субстрат целевой PRTase, может быть летальным для клеток, когда основание включено в предшественники нуклеотидов, тогда как на мутант с нефункциональной PRTase меньше влияет токсичность аналога основания. Подход встречного отбора включал использование вектора интеграции, который обеспечивает эктопическую экспрессию гена, кодирующего PRTase, в фоновом хозяине, дефектном по PRTase, для нацеленных на ген делеций. Первоначально выбирают интеграцию рекомбинантной плазмиды с одним кроссовером, и событие рекомбинации делает хозяина чувствительным к PRTase-специфическому основанию аналога из-за экспрессии гена PRTase, переносимого плазмидой. Вырезание интегрированной плазмиды после второго события рекомбинации восстанавливает фенотип устойчивости к основному аналогу хозяина, который затем служит в качестве стратегии встречного отбора для быстрой идентификации свободных от плазмид рекомбинантов, которые несут либо аллель дикого типа, либо мутантный аллель целевой ген.Подход встречного отбора на основе upp ранее использовался для замены аллелей в B. subtilis и Enterococcus faecalis (24, 33).
Первоначально выбирают интеграцию рекомбинантной плазмиды с одним кроссовером, и событие рекомбинации делает хозяина чувствительным к PRTase-специфическому основанию аналога из-за экспрессии гена PRTase, переносимого плазмидой. Вырезание интегрированной плазмиды после второго события рекомбинации восстанавливает фенотип устойчивости к основному аналогу хозяина, который затем служит в качестве стратегии встречного отбора для быстрой идентификации свободных от плазмид рекомбинантов, которые несут либо аллель дикого типа, либо мутантный аллель целевой ген.Подход встречного отбора на основе upp ранее использовался для замены аллелей в B. subtilis и Enterococcus faecalis (24, 33).
В настоящем исследовании мы сообщаем о разработке улучшенной безмаркерной системы замены генов для L. acidophilus NCFM, которая включает использование upp в качестве контрселективного маркера для существующей системы нокаута на основе pORI для положительного отбора двойных рекомбинанты. Мы продемонстрировали эффективность схемы контрселекции upp , удалив ген slpX (LBA0512), кодирующий новый секретируемый белок, который, как было обнаружено, связан с комплексом S-слоя в L.ацидофилус . Это исследование продемонстрировало первую функциональную контрселективную систему замены генов для рода Lactobacillus .
Мы продемонстрировали эффективность схемы контрселекции upp , удалив ген slpX (LBA0512), кодирующий новый секретируемый белок, который, как было обнаружено, связан с комплексом S-слоя в L.ацидофилус . Это исследование продемонстрировало первую функциональную контрселективную систему замены генов для рода Lactobacillus .
МАТЕРИАЛЫ И МЕТОДЫ
Штаммы бактерий и условия роста. Штаммы бактерий и плазмиды, использованные в этом исследовании, показаны в таблице 1. штамм L. acidophilus размножали в бульоне MRS (Difco Laboratories, Inc., Детройт, Мичиган) статически в условиях окружающей атмосферы или на твердой среде MRS с добавлением 1.5% (вес / объем) агар (Difco) в анаэробных условиях при 37 ° C или 42 ° C, как указано ниже. Рекомбинантные штаммы отбирали в присутствии 2 мкг / мл эритромицина (Em) (Sigma-Aldrich, Сент-Луис, Миссури) и / или 5 мкг / мл хлорамфеникола (Cm) (Sigma), когда это необходимо. Для контрселекции свободных от плазмид двойных рекомбинантов, 5-фторурацил (5-FU) (Sigma), аналог урацила, был добавлен к предварительно стерилизованной полуопределенной агаризованной среде (SDM) (30) в конечной концентрации 100 мкг / мл из 60 -мг / мл маточный раствор, приготовленный с диметилсульфоксидом (ДМСО) (Sigma), как описано Kristich et al.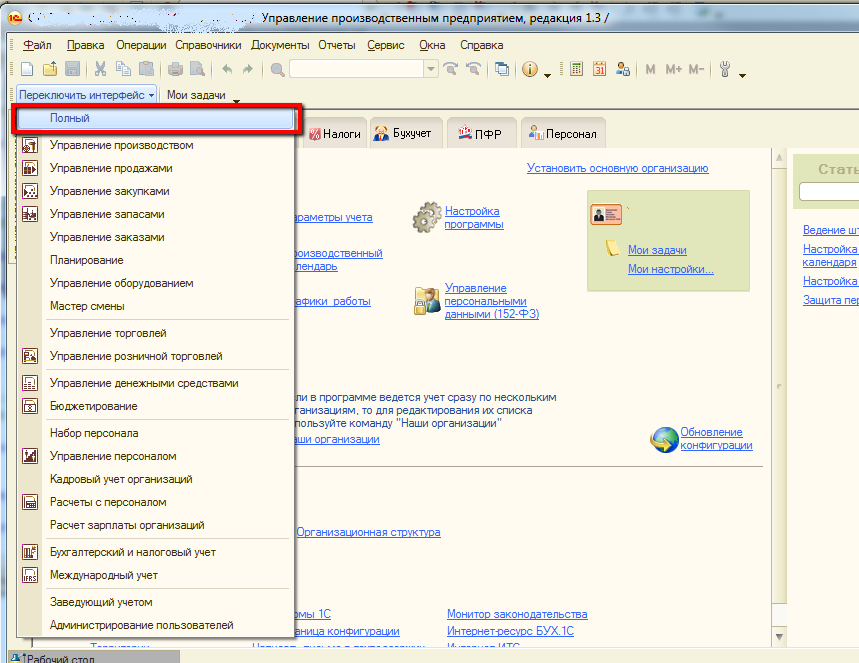
Бактериальные штаммы и плазмиды, использованные в этом исследовании
Штаммы Escherichia coli выращивали в среде для инфузии мозга и сердца (BHI) (Difco) при 37 ° C с аэрацией. E. coli EC101 размножали в присутствии 40 мкг / мл канамицина. При необходимости добавляли Em и Cm до конечных концентраций 200 мкг / мл и 15 мкг / мл соответственно.
Выделение ДНК и манипуляции. Геномную ДНК L. acidophilus NCFM выделяли методом, описанным Уокером и Клаенхаммером (52), или с использованием набора для выделения микробной ДНК Mo Bio UltraClean (Mo Bio Laboratories, Карлсбад, Калифорния).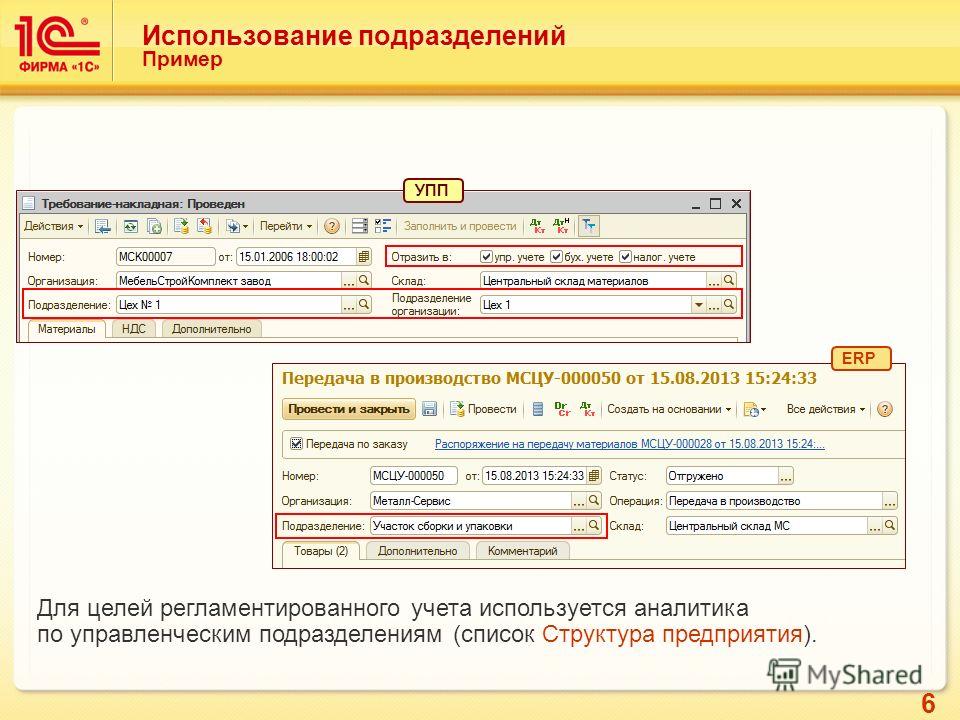 Плазмидную ДНК из E. coli выделяли с использованием набора QIAprep Spin miniprep (Qiagen Inc., Валенсия, Калифорния). Рестрикционные ферменты (Roche Molecular Biochemicals, Indianapolis, IN) использовали в соответствии с инструкциями, предоставленными производителем. Лигирование ДНК выполняли с использованием ДНК-лигазы Т4 (New England Biolabs, Беверли, Массачусетс) или набора для лигирования ДНК Fast-Link (Epicenter Biotechnologies, Мэдисон, Висконсин) в соответствии с рекомендациями производителей. Праймеры для ПЦР (см. Таблицу S1 в дополнительном материале) были синтезированы Integrated DNA Technologies (Coralville, IA). Для клонирования ампликоны ПЦР были созданы с использованием ДНК-полимеразы PfuUltra II Fusion HS (Stratagene Corp., La Jolla, CA) в соответствии с инструкциями поставщика. Обычные ПЦР-амплификации для скрининга рекомбинантов выполняли с использованием стандартных протоколов и ДНК-полимеразы Choice- Taq Blue (Denville Scientific Inc., Метучен, Нью-Джерси).
Плазмидную ДНК из E. coli выделяли с использованием набора QIAprep Spin miniprep (Qiagen Inc., Валенсия, Калифорния). Рестрикционные ферменты (Roche Molecular Biochemicals, Indianapolis, IN) использовали в соответствии с инструкциями, предоставленными производителем. Лигирование ДНК выполняли с использованием ДНК-лигазы Т4 (New England Biolabs, Беверли, Массачусетс) или набора для лигирования ДНК Fast-Link (Epicenter Biotechnologies, Мэдисон, Висконсин) в соответствии с рекомендациями производителей. Праймеры для ПЦР (см. Таблицу S1 в дополнительном материале) были синтезированы Integrated DNA Technologies (Coralville, IA). Для клонирования ампликоны ПЦР были созданы с использованием ДНК-полимеразы PfuUltra II Fusion HS (Stratagene Corp., La Jolla, CA) в соответствии с инструкциями поставщика. Обычные ПЦР-амплификации для скрининга рекомбинантов выполняли с использованием стандартных протоколов и ДНК-полимеразы Choice- Taq Blue (Denville Scientific Inc., Метучен, Нью-Джерси). Продукты ПЦР анализировали на 0,8% агарозных гелях и очищали с использованием набора для выделения ДНК из геля Zymoclean (Zymo Research Corp., Orange, CA) или набора для экстракции из геля QIAquick (Qiagen). Секвенирование ДНК было выполнено Davis Sequencing, Inc. (Дэвис, Калифорния).
Продукты ПЦР анализировали на 0,8% агарозных гелях и очищали с использованием набора для выделения ДНК из геля Zymoclean (Zymo Research Corp., Orange, CA) или набора для экстракции из геля QIAquick (Qiagen). Секвенирование ДНК было выполнено Davis Sequencing, Inc. (Дэвис, Калифорния).
E.coli были получены химически компетентные клетки и трансформированы в соответствии с процедурами, ранее описанными Hanahan (27). Клетки L. acidophilus получали для электротрансформации, по существу, как описано Walker et al. (51) со следующими модификациями, адаптированными из процедуры Wei et al. (55). Вкратце, клетки стационарной фазы инокулировали в бульон MRS (2% инокулята) и выращивали в течение 3 часов (OD 600 , ~ 0,1-0,2). Затем добавляли стерилизованный фильтром маточный раствор пенициллина G для получения конечной концентрации 10 мкг / мл, и культуру инкубировали в течение 1.За 5–2,0 ч до сбора урожая.
Конструирование мутанта с делецией upp . Чтобы сконструировать изогенный мутант Δ upp с внутренней делецией 315 п.н. в гене upp (LBA0770), сначала нужно выделить вышестоящий сегмент хромосомной ДНК (596 п.н.) и нижестоящий сегмент хромосомной ДНК (610 п.н.), фланкирующий область делеции, амплифицировали с помощью ПЦР с парами праймеров upp1U-F / upp2U-R и upp3D-F / upp4D-R (см. Таблицу S1 в дополнительном материале) соответственно. Два ПЦР-фрагмента очищали в геле и соединяли с помощью сплайсинга методом ПЦР с удлинением перекрытия (SOE-PCR) (28), где эквимолярные концентрации каждого продукта ПЦР (от 10 до 15 нг) объединяли и использовали в качестве матриц для амплификации в 50 мкл смеси для ПЦР с праймерами upp1U-F и upp4D-R.Все ПЦР проводили с 25-30 циклами амплификации, чтобы минимизировать накопление ошибок неправильного включения нуклеотидов в продукты ПЦР. Полученные продукты SOE-PCR очищали на геле, рестриктировали с помощью XbaI и SacI, лигировали в аналогично расщепленный вектор pORI19 и трансформировали в E.
Чтобы сконструировать изогенный мутант Δ upp с внутренней делецией 315 п.н. в гене upp (LBA0770), сначала нужно выделить вышестоящий сегмент хромосомной ДНК (596 п.н.) и нижестоящий сегмент хромосомной ДНК (610 п.н.), фланкирующий область делеции, амплифицировали с помощью ПЦР с парами праймеров upp1U-F / upp2U-R и upp3D-F / upp4D-R (см. Таблицу S1 в дополнительном материале) соответственно. Два ПЦР-фрагмента очищали в геле и соединяли с помощью сплайсинга методом ПЦР с удлинением перекрытия (SOE-PCR) (28), где эквимолярные концентрации каждого продукта ПЦР (от 10 до 15 нг) объединяли и использовали в качестве матриц для амплификации в 50 мкл смеси для ПЦР с праймерами upp1U-F и upp4D-R.Все ПЦР проводили с 25-30 циклами амплификации, чтобы минимизировать накопление ошибок неправильного включения нуклеотидов в продукты ПЦР. Полученные продукты SOE-PCR очищали на геле, рестриктировали с помощью XbaI и SacI, лигировали в аналогично расщепленный вектор pORI19 и трансформировали в E. coli Dh20B, несущую плазмиду-помощник pTRK669 (NCK1391), с отбором на среде BHI, содержащей как Em, так и Cm. . Полученную рекомбинантную плазмиду pTRK934 электропорировали в L. acidophilus NCK1392.Процедуры, используемые для выделения свободных от плазмид рекомбинантов после хромосомной интеграции pTRK934 и рекомбинации с двойным кроссинговером, выполняли в основном, как описано ранее (19, 46). Делеции в рамке считывания в мутантах Δ upp подтверждали секвенированием ДНК продуктов ПЦР, полученных с парой праймеров upp-up / upp-dw (см. Таблицу S1 в дополнительном материале), специфичных для области, фланкирующей мишень для делеции. Анализ
coli Dh20B, несущую плазмиду-помощник pTRK669 (NCK1391), с отбором на среде BHI, содержащей как Em, так и Cm. . Полученную рекомбинантную плазмиду pTRK934 электропорировали в L. acidophilus NCK1392.Процедуры, используемые для выделения свободных от плазмид рекомбинантов после хромосомной интеграции pTRK934 и рекомбинации с двойным кроссинговером, выполняли в основном, как описано ранее (19, 46). Делеции в рамке считывания в мутантах Δ upp подтверждали секвенированием ДНК продуктов ПЦР, полученных с парой праймеров upp-up / upp-dw (см. Таблицу S1 в дополнительном материале), специфичных для области, фланкирующей мишень для делеции. Анализ
PM. Эксперименты с фенотипическими микрочипами (PM) были выполнены Biolog Inc.(Хейворд, Калифорния). Технология PM основана на химической реакции между бактериями и тетразолиевым красителем. Если среда в лунке поддерживает рост бактерий, метаболизирующие клетки уменьшают количество тетразолия и дают цвет, который можно измерить с помощью прибора OmniLog в течение 24 часов. Информация о посевной среде и концентрации субстратов является частной собственностью (www.biolog.com
). В этом исследовании использовались двенадцать 96-луночных PM (от PM09 до PM20). Эти массивы использовались для следующих анализов: PM09, осмолиты; PM10, pH; и от PM11 до PM20, химическая чувствительность (см. таблицы S2 и S3 в дополнительном материале).Ночные культуры штаммов L. acidophilus ресуспендировали в бульоне MRS в соответствии с процедурами Biolog (www.biolog.com
) и добавляли в каждую лунку 12 96-луночных планшетов. При отрицательных реакциях цвет образуется незначительно или совсем отсутствует, тогда как цвет образуется при положительных реакциях. Инструмент OmniLog захватил цифровые изображения PM, файлы были впоследствии отображены в виде кинетических графиков для дыхания и роста, а программное обеспечение использовалось для расчета площади под кривой для каждого штамма.
Информация о посевной среде и концентрации субстратов является частной собственностью (www.biolog.com
). В этом исследовании использовались двенадцать 96-луночных PM (от PM09 до PM20). Эти массивы использовались для следующих анализов: PM09, осмолиты; PM10, pH; и от PM11 до PM20, химическая чувствительность (см. таблицы S2 и S3 в дополнительном материале).Ночные культуры штаммов L. acidophilus ресуспендировали в бульоне MRS в соответствии с процедурами Biolog (www.biolog.com
) и добавляли в каждую лунку 12 96-луночных планшетов. При отрицательных реакциях цвет образуется незначительно или совсем отсутствует, тогда как цвет образуется при положительных реакциях. Инструмент OmniLog захватил цифровые изображения PM, файлы были впоследствии отображены в виде кинетических графиков для дыхания и роста, а программное обеспечение использовалось для расчета площади под кривой для каждого штамма.
Исследование экспрессии генов на микрочипах. (i) Создание ДНК-олигомассивов NCFM L. acidophilus . OligoArray 2.1 (45) использовали для конструирования олигонуклеотидов для конструирования олигонуклеотидных микрочипов NCFM (олигонуклеотидов). Диапазон используемых длин олигонуклеотидов составлял от 60 до 70 нуклеотидов, используемый диапазон температур плавления составлял от 80 до 96 ° C, а используемый диапазон содержания G + C составлял от 25 до 50% в зависимости от содержания G + C в геноме NCFM (34,71 %). Пороговые значения для отклонения олигонуклеотидов, которые могут образовывать стабильные вторичные структуры и возможной перекрестной гибридизации, составляли 65 ° C.Олигонуклеотиды, содержащие гомополимеры AAAAA, TTTTT, GGGGG, CCCCC или более длинные, были отклонены. Олигонуклеотиды были синтезированы Integrated DNA Technologies и нанесены на предметные стекла. Каждый олигонуклеотид (представляющий каждый ген) был нанесен шесть раз случайным образом.
acidophilus . OligoArray 2.1 (45) использовали для конструирования олигонуклеотидов для конструирования олигонуклеотидных микрочипов NCFM (олигонуклеотидов). Диапазон используемых длин олигонуклеотидов составлял от 60 до 70 нуклеотидов, используемый диапазон температур плавления составлял от 80 до 96 ° C, а используемый диапазон содержания G + C составлял от 25 до 50% в зависимости от содержания G + C в геноме NCFM (34,71 %). Пороговые значения для отклонения олигонуклеотидов, которые могут образовывать стабильные вторичные структуры и возможной перекрестной гибридизации, составляли 65 ° C.Олигонуклеотиды, содержащие гомополимеры AAAAA, TTTTT, GGGGG, CCCCC или более длинные, были отклонены. Олигонуклеотиды были синтезированы Integrated DNA Technologies и нанесены на предметные стекла. Каждый олигонуклеотид (представляющий каждый ген) был нанесен шесть раз случайным образом.
(ii) Условия эксперимента и выделение РНК. Штамм NCFM дикого типа и изогенный мутант Δ upp выращивали в свежей среде MRS с использованием 2% инокулята из ночных культур. Аликвоты клеток собирали при OD 600 , равном ∼0.3 (ранняя логарифмическая фаза) и 0,8 (средняя логарифмическая фаза) быстрым центрифугированием при комнатной температуре. Осадки клеток мгновенно замораживали на бане с сухим льдом и этанолом и хранили при -80 ° C. Образцы были собраны из двух независимых экспериментальных повторностей (представляющих две биологические повторности). Выделение общей РНК, обработку ДНКазой и очистку РНК выполняли в основном, как описано ранее (26). Отсутствие геномной ДНК в очищенных образцах РНК подтверждали проведением ПЦР с праймерами, специфичными для генов NCFM.
Аликвоты клеток собирали при OD 600 , равном ∼0.3 (ранняя логарифмическая фаза) и 0,8 (средняя логарифмическая фаза) быстрым центрифугированием при комнатной температуре. Осадки клеток мгновенно замораживали на бане с сухим льдом и этанолом и хранили при -80 ° C. Образцы были собраны из двух независимых экспериментальных повторностей (представляющих две биологические повторности). Выделение общей РНК, обработку ДНКазой и очистку РНК выполняли в основном, как описано ранее (26). Отсутствие геномной ДНК в очищенных образцах РНК подтверждали проведением ПЦР с праймерами, специфичными для генов NCFM.
(iii) Синтез зонда кДНК и гибридизация олигомассивов. Обратную транскрипцию и флуоресцентное мечение идентичных количеств (20 мкг) очищенной РНК проводили с использованием системы непрямого мечения кДНК SuperScript для микроматриц ДНК (Invitrogen, Карлсбад, Калифорния) в соответствии с производителя, за исключением того, что 6 мкг случайных праймеров (Invitrogen) использовали для каждых 30 мкл реакционной смеси синтеза первой цепи кДНК. Меченые образцы кДНК, представляющие мутантные штаммы NCFM и Δ upp , были соединены с монореактивными красителями Cy3 и Cy5 (GE Healthcare Bio-Sciences Corp., Пискатауэй, штат Нью-Джерси) соответственно. Сравнительную гибридизацию проводили с образцами, представляющими как NCFM, так и производное Δ upp на одних и тех же фазах роста. Перед гибридизацией слайды олигомассивов предварительно гибридизовали в буфере, содержащем 0,1% (вес / объем) додецилсульфата натрия (SDS) (Invitrogen), 0,1% (вес / объем) бычьего сывороточного альбумина (Invitrogen) и 5 × SSC (1 × SSC представляет собой 0,15 М NaCl плюс 0,015 М цитрат натрия; Invitrogen) при 42 ° C в течение 1 ч, а затем промывают, погружая их пять раз в стерильную дистиллированную воду и дважды в изопропанол и сушат вращением.Попарно меченные Cy3 и Cy5 зонды сушили с помощью вакуумной системы Vacufuge speed (Epperdorf, Westbury, NY) и ресуспендировали в 48-50 мкл (общий объем) гибридизационного буфера, содержащего 5 × SSC, 5 × раствор Денхардта (Invitrogen).
Меченые образцы кДНК, представляющие мутантные штаммы NCFM и Δ upp , были соединены с монореактивными красителями Cy3 и Cy5 (GE Healthcare Bio-Sciences Corp., Пискатауэй, штат Нью-Джерси) соответственно. Сравнительную гибридизацию проводили с образцами, представляющими как NCFM, так и производное Δ upp на одних и тех же фазах роста. Перед гибридизацией слайды олигомассивов предварительно гибридизовали в буфере, содержащем 0,1% (вес / объем) додецилсульфата натрия (SDS) (Invitrogen), 0,1% (вес / объем) бычьего сывороточного альбумина (Invitrogen) и 5 × SSC (1 × SSC представляет собой 0,15 М NaCl плюс 0,015 М цитрат натрия; Invitrogen) при 42 ° C в течение 1 ч, а затем промывают, погружая их пять раз в стерильную дистиллированную воду и дважды в изопропанол и сушат вращением.Попарно меченные Cy3 и Cy5 зонды сушили с помощью вакуумной системы Vacufuge speed (Epperdorf, Westbury, NY) и ресуспендировали в 48-50 мкл (общий объем) гибридизационного буфера, содержащего 5 × SSC, 5 × раствор Денхардта (Invitrogen).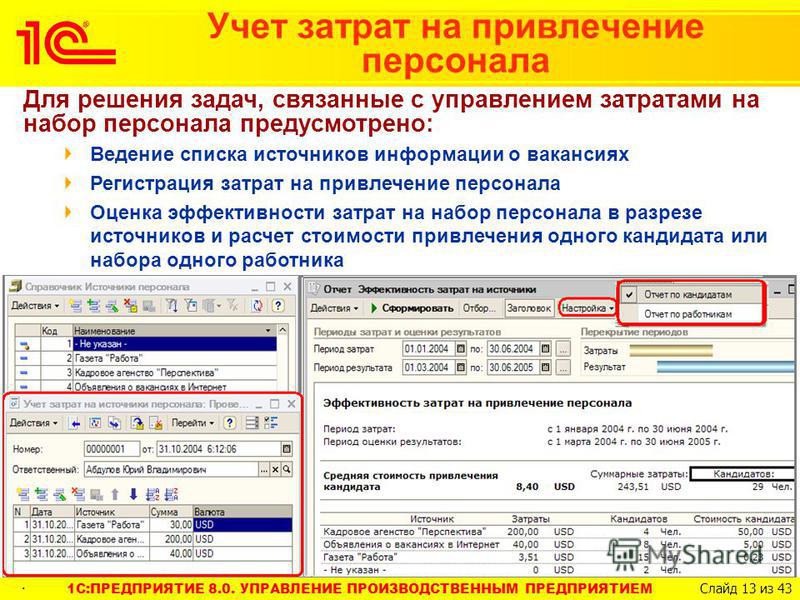 30% (об. / Об.) Формамида, 0,1% SDS и 200 мкг / мл ДНК спермы лосося (Invitrogen). Полученные смеси зондов инкубировали при 95 ° C в течение 2 минут и гибридизовали с олигоанализаторами в течение 18-20 часов при 42 ° C. Матрицы после гибридизации сначала промывали предварительно нагретым (42 ° C) промывочным буфером, содержащим 1 × SSC и 0.03% SDS, а затем в промывочном буфере, содержащем 0,5 × SSC и 0,03% SDS, при комнатной температуре. Затем матрицы промывали 0,2 × SSC, после чего следовала заключительная промывка 0,05 × SSC. Каждый этап промывки длился 5 мин.
30% (об. / Об.) Формамида, 0,1% SDS и 200 мкг / мл ДНК спермы лосося (Invitrogen). Полученные смеси зондов инкубировали при 95 ° C в течение 2 минут и гибридизовали с олигоанализаторами в течение 18-20 часов при 42 ° C. Матрицы после гибридизации сначала промывали предварительно нагретым (42 ° C) промывочным буфером, содержащим 1 × SSC и 0.03% SDS, а затем в промывочном буфере, содержащем 0,5 × SSC и 0,03% SDS, при комнатной температуре. Затем матрицы промывали 0,2 × SSC, после чего следовала заключительная промывка 0,05 × SSC. Каждый этап промывки длился 5 мин.
(iv) Сбор и анализ данных. Интенсивность флуоресценции регистрировали при разрешении 10 мкм на пиксель с помощью сканера микрочипов ScanArray 4000 (Packard Biochip BioScience; Biochip Technologies LLC) и сохраняли в виде файлов изображений TIFF. Была произведена количественная оценка интенсивности сигнала, вычиталось значение фона и данные были нормализованы с использованием QuantArray 3.0 (Packard Bioscience). Пятна анализировали с помощью адаптивного количественного анализа. Данные были нормализованы по медиане (7). Среднее соотношение Cy5 / Cy3 для 12 пятен для каждого гена (6 репликных пятен на слайд на биологический повтор) было определено и представляло изменение уровней экспрессии гена в мутанте Δ upp (Cy5) по сравнению со штаммом NCFM (Cy3). . Доверительные интервалы и значения P для изменения также были рассчитаны с использованием двухвыборочного теста t .Значения P ≤0,05 считались значимыми.
Данные были нормализованы по медиане (7). Среднее соотношение Cy5 / Cy3 для 12 пятен для каждого гена (6 репликных пятен на слайд на биологический повтор) было определено и представляло изменение уровней экспрессии гена в мутанте Δ upp (Cy5) по сравнению со штаммом NCFM (Cy3). . Доверительные интервалы и значения P для изменения также были рассчитаны с использованием двухвыборочного теста t .Значения P ≤0,05 считались значимыми.
Конструирование вектора интеграции с возможностью обратного выбора. Для создания вектора интеграции с возможностью обратного выбора на основе pORI сайт множественного клонирования (MCS) вектора pORI28 был заменен кассетой экспрессии upp (P- upp ) и ген lacZ ‘( lacZ -альфа); Ген lacZ ‘был использован для сине-белого скрининга трансформантов E. coli посредством альфа-комплементации Δ lacZ ‘ E.coli клонирующие хозяева (рис. 1). Сначала ДНК плазмиды pORI28 расщепляли BglII и XbaI для удаления области MCS. Затем 5′-выступы рестриктированной плазмиды pORI28 заполняли с использованием ДНК-полимеразы PfuUltra II Fusion HS на основе протокола набора для полировки Stratagene PCR (Stratagene). Затем ген lacZ ‘с его промотором и сайтом связывания белка-активатора катаболита (CAP) амплифицировали из вектора pUC19 с парой праймеров pUC19lacZ-F / pUC19lacZ-R (см. Таблицу S1 в дополнительном материале) с использованием 10 нг ДНК вектора pUC19 в качестве матрицы для амплификации в 50 мкл реакционной смеси.Ген L. acidophilus NCFM upp с его предполагаемой промоторной областью амплифицировали с помощью ПЦР с парой праймеров upp-F / upp-R (см. Таблицу S1 в дополнительном материале). Оба очищенных ПЦР-ампликона были соединены с помощью SOE-PCR, как описано выше, с использованием пары праймеров pUC19lacZ-F / upp-R. Полученные продукты слияния lacZ ‘-P- upp лигировали в остов pORI28 с тупыми концами и трансформировали в E.coli NCK1391, и трансформанты отбирали на чашках со средой BHI, содержащих Em и Cm в присутствии X -Гал (5-бром-4-хлор-3-индолил-β-d-галактопиранозид) и изопропил-β-d-тиогалактопиранозид (IPTG).
Затем 5′-выступы рестриктированной плазмиды pORI28 заполняли с использованием ДНК-полимеразы PfuUltra II Fusion HS на основе протокола набора для полировки Stratagene PCR (Stratagene). Затем ген lacZ ‘с его промотором и сайтом связывания белка-активатора катаболита (CAP) амплифицировали из вектора pUC19 с парой праймеров pUC19lacZ-F / pUC19lacZ-R (см. Таблицу S1 в дополнительном материале) с использованием 10 нг ДНК вектора pUC19 в качестве матрицы для амплификации в 50 мкл реакционной смеси.Ген L. acidophilus NCFM upp с его предполагаемой промоторной областью амплифицировали с помощью ПЦР с парой праймеров upp-F / upp-R (см. Таблицу S1 в дополнительном материале). Оба очищенных ПЦР-ампликона были соединены с помощью SOE-PCR, как описано выше, с использованием пары праймеров pUC19lacZ-F / upp-R. Полученные продукты слияния lacZ ‘-P- upp лигировали в остов pORI28 с тупыми концами и трансформировали в E.coli NCK1391, и трансформанты отбирали на чашках со средой BHI, содержащих Em и Cm в присутствии X -Гал (5-бром-4-хлор-3-индолил-β-d-галактопиранозид) и изопропил-β-d-тиогалактопиранозид (IPTG). Положительные клоны выглядели как синие колонии из-за экспрессии гена lacZ ‘в рекомбинантной плазмиде. Полученный в результате противоселективный вектор интеграции, pTRK935, подвергали секвенированию ДНК для подтверждения целостности P- upp и MCS в гене lacZ ‘.
Положительные клоны выглядели как синие колонии из-за экспрессии гена lacZ ‘в рекомбинантной плазмиде. Полученный в результате противоселективный вектор интеграции, pTRK935, подвергали секвенированию ДНК для подтверждения целостности P- upp и MCS в гене lacZ ‘.
Построение вектора интегрирования с возможностью выбора счетчика pTRK935. ori , ориджин репликации pWV01; erm , ген, кодирующий устойчивость к Em; lacZ ‘, lacZ -alpha ген из pUC19; MCS, MCS из pUC19.Показаны только уникальные сайты ограничения.
Конструирование мутанта Δ slpX с использованием контрселективной системы замены гена. Делецию в рамке считывания длиной 1356 п.о. в гене slpX (LBA0512) конструировали путем первой амплификации сегментов ДНК длиной 767 п.о. и 747 п.о., фланкирующих восходящий поток. и нижележащие области мишени для делеции, соответственно, с использованием пар праймеров slpX1U-F / slpX2U-R и slpX3D-F / slpX4D-R (см. Таблицу S1 в дополнительном материале). Два очищенных продукта ПЦР были слиты и амплифицированы для создания копий аллеля Δ slpX методом SOE-PCR, как описано выше, расщеплены BamHI и SacI и лигированы в вектор интеграции pTRK935 с совместимыми концами.Смесь для лигирования трансформировали в E. coli EC101 с селекцией на среде BHI, содержащей канамицин, Em, X-Gal и IPTG. Полученную плазмиду, несущую аллель Δ slpX , pTRK936, электропорировали в штамм-хозяин L. acidophilus Δ upp , несущий pTRK669, NCK1910 (таблица 1). Один трансформант Em r Cm r , несущий обе плазмиды, выращивали в течение ночи в среде MRS, содержащей 2 мкг / мл Em и 2 мкг / мл Cm, и переносили три раза (1% инокулята) в среду MRS с Em (приблизительно.30 поколений) на водяной бане 42 ° C. Хромосомные интегранты отбирали путем посева реплик образцов на агар MRS с добавлением Em или Cm. Отбирали один интегрант Em r Cm s и размножали в среде MRS с Em при 37 ° C в течение ночи, после чего следовало от трех до пяти переносов (1% инокулята) в среду MRS без Em (примерно от 30 до 50 поколений).
Таблицу S1 в дополнительном материале). Два очищенных продукта ПЦР были слиты и амплифицированы для создания копий аллеля Δ slpX методом SOE-PCR, как описано выше, расщеплены BamHI и SacI и лигированы в вектор интеграции pTRK935 с совместимыми концами.Смесь для лигирования трансформировали в E. coli EC101 с селекцией на среде BHI, содержащей канамицин, Em, X-Gal и IPTG. Полученную плазмиду, несущую аллель Δ slpX , pTRK936, электропорировали в штамм-хозяин L. acidophilus Δ upp , несущий pTRK669, NCK1910 (таблица 1). Один трансформант Em r Cm r , несущий обе плазмиды, выращивали в течение ночи в среде MRS, содержащей 2 мкг / мл Em и 2 мкг / мл Cm, и переносили три раза (1% инокулята) в среду MRS с Em (приблизительно.30 поколений) на водяной бане 42 ° C. Хромосомные интегранты отбирали путем посева реплик образцов на агар MRS с добавлением Em или Cm. Отбирали один интегрант Em r Cm s и размножали в среде MRS с Em при 37 ° C в течение ночи, после чего следовало от трех до пяти переносов (1% инокулята) в среду MRS без Em (примерно от 30 до 50 поколений). ). Культуры были серийно разведены, и от 10 -3 до 10 -4 разведений высевали на планшеты SDM с добавлением 5-ФУ и инкубировали при 37 ° C в анаэробных условиях в течение 48-72 ч до появления видимых колоний.Восемь колоний 5-FU r были отобраны для ПЦР-анализа области slpX с использованием праймеров slpX-up и slpX-dw (см. Таблицу S1 в дополнительном материале), которые специфически отжигаются с фланкирующей областью slpX . Делеции в рамке считывания подтверждали секвенированием ДНК, как описано выше.
). Культуры были серийно разведены, и от 10 -3 до 10 -4 разведений высевали на планшеты SDM с добавлением 5-ФУ и инкубировали при 37 ° C в анаэробных условиях в течение 48-72 ч до появления видимых колоний.Восемь колоний 5-FU r были отобраны для ПЦР-анализа области slpX с использованием праймеров slpX-up и slpX-dw (см. Таблицу S1 в дополнительном материале), которые специфически отжигаются с фланкирующей областью slpX . Делеции в рамке считывания подтверждали секвенированием ДНК, как описано выше.
Экстракция и очистка белков S-слоя. Культуры размножали в течение ночи в 100 мл среды MRS, и клетки собирали центрифугированием при 8000 × g в течение 20 минут при 4 ° C.Сухой вес осадка клеток из 100-мл культур составлял от 600 до 700 мг, что соответствует влажному весу от 5,4 до 6,3 г. После одной промывки холодной дистиллированной водой гранулы суспендировали в 5 M LiCl при соотношении 1 мл LiCl к 10 мг биомассы и инкубировали в течение 15 мин при 4 ° C при легком перемешивании.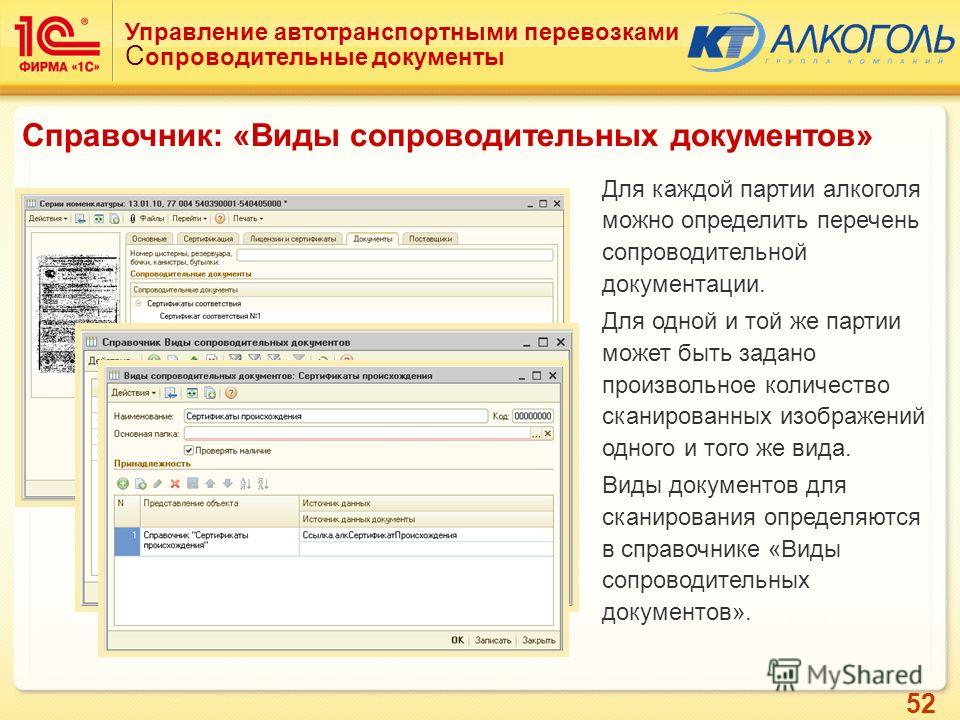 После центрифугирования супернатант (~ 60 мл) удаляли и диализовали в течение 24 часов против дистиллированной воды при 4 ° C с использованием мембраны Spectrapor 6-8000. Белый осадок, образовавшийся во время диализа, собирали центрифугированием при 40000 × g в течение 20 минут при 4 ° C.Для удаления белков, соосажденных с белком S-слоя, осадок ресуспендировали в 1 М LiCl и перемешивали в течение 15 мин при 4 ° C. После центрифугирования при 40000 × г в течение 20 минут при 4 ° C осадок промывали 15 мл холодной дистиллированной воды. Затем каждый осадок разделяли на две части, одну из которых использовали для прямого масс-спектрометрического (МС) анализа, а другую использовали для трипсинолиза и анализа лазерной десорбционной ионизации с использованием матрицы.
После центрифугирования супернатант (~ 60 мл) удаляли и диализовали в течение 24 часов против дистиллированной воды при 4 ° C с использованием мембраны Spectrapor 6-8000. Белый осадок, образовавшийся во время диализа, собирали центрифугированием при 40000 × g в течение 20 минут при 4 ° C.Для удаления белков, соосажденных с белком S-слоя, осадок ресуспендировали в 1 М LiCl и перемешивали в течение 15 мин при 4 ° C. После центрифугирования при 40000 × г в течение 20 минут при 4 ° C осадок промывали 15 мл холодной дистиллированной воды. Затем каждый осадок разделяли на две части, одну из которых использовали для прямого масс-спектрометрического (МС) анализа, а другую использовали для трипсинолиза и анализа лазерной десорбционной ионизации с использованием матрицы.
MS-анализ чистых белков S-слоя до и после трипсинолиза.Для прямого МС-анализа чистые белки S-слоя ресуспендировали в растворе, содержащем 50% (об. / Об.) Ацетонитрила и 4% (об. / Об.) Муравьиной кислоты в дистиллированной воде (общий объем, 500 мкл) в течение ночи при 4 ° C. Затем добавляли 200 мкл того же раствора ацетонитрил-муравьиной кислоты и смесь интенсивно перемешивали. Полная ресолюбилизация белкового осадка S-слоя никогда не наблюдалась; поэтому суспензию центрифугировали (12000 × г, , 10 мин, 4 ° C), супернатант удаляли и использовали для анализа МС.
Затем добавляли 200 мкл того же раствора ацетонитрил-муравьиной кислоты и смесь интенсивно перемешивали. Полная ресолюбилизация белкового осадка S-слоя никогда не наблюдалась; поэтому суспензию центрифугировали (12000 × г, , 10 мин, 4 ° C), супернатант удаляли и использовали для анализа МС.
Для снятия пептидных фингерпринтов чистые белки S-слоя (хранящиеся при -20 ° C) подвергали трипсинолизу. Расщепление трипсином S-слоя выполняли в течение ночи при 37 ° C и останавливали трифторуксусной кислотой (Sigma-Aldrich, St. Quentin-Fallavier, Франция). Затем супернатанты, содержащие пептиды, сушили с использованием вакуумной системы Speed и хранили при -20 ° C до проведения анализа масс-спектрометрии.
MS. Все масс-спектры были получены с использованием гибридного квадрупольного времяпролетного масс-спектрометра (QStar XL; MDS Sciex, Торонто, Канада).Для экспериментов по лазерной десорбционной ионизации с использованием матрицы образцы ионизировали лазерным лучом (λ = 337 нм, 20 Гц), и фрагмент пептида β-казеина от остатка 193 до остатка 209 использовали в качестве калибровочного стандарта. Более характерные однозарядные ионы подвергались фрагментации с энергией столкновения от 50 до 90 кэВ.
Более характерные однозарядные ионы подвергались фрагментации с энергией столкновения от 50 до 90 кэВ.
Для экспериментов по ионизации электрораспылением к масс-спектрометру QStar XL подключили систему жидкостной нано-жидкостной хроматографии (LC-насадки; Dionex Voisins, Le Bretonneux, Франция).Масс-спектрометр работал в режиме положительных ионов. К источнику микроионов было приложено напряжение около 5 кВ. Данные МС и тандемной МС (МС / МС) получали в непрерывном режиме. Прибор был откалиброван с помощью многоточечной калибровки с использованием фрагментных ионов, которые возникли в результате индуцированного столкновением разложения фрагмента пептида β-казеина от остатка 193 до остатка 209. Для выполнения анализа МС / МС с двух- и трехзарядным предшественником использовался анализ, управляемый данными. ионы. МС / МС спектры были получены от m / z 50 до m / z 2000.Масс-спектрометр работал в режиме зависимости от данных автоматически с переключением между МС и МС / МС с использованием программного обеспечения Analyst QS 1. 1. Затем все данные (MS и MS / MS) были проанализированы с помощью MASCOT (v.2.1) для поиска в нескольких базах данных (MSDB и Интернет-база данных под названием «UMR») для идентификации белков, присутствующих в образцах.
1. Затем все данные (MS и MS / MS) были проанализированы с помощью MASCOT (v.2.1) для поиска в нескольких базах данных (MSDB и Интернет-база данных под названием «UMR») для идентификации белков, присутствующих в образцах.
Анализы с нагрузкой на стресс. Штаммы выращивали в бульоне MRS до OD 600 от 0,25 до 0,3 (ранняя логарифмическая фаза) перед тем, как подвергать их использованным нагрузкам (7).Культуры делили на порции равного объема и собирали при комнатной температуре. Клетки ресуспендировали в равных объемах (i) бульона MRS с 0,3% (мас. / Об.) Свиной желчи (Sigma), (ii) бульона MRS с 2,5% (мас. / Об.) Бычьей желчи (Difco), (iii) бульона MRS с 10% (мас. / Об.) NaCl, (iv) бульон MRS с 15% (об. / Об.) Этанолом и (v) подкисленный бульон MRS при pH 3,5 (отрегулированный с помощью молочной кислоты) и pH 3,0 (отрегулированный с помощью HCl). Подсчет жизнеспособных клеток определяли через 2 часа инкубации при 37 ° C путем посева на среду MRS.Для анализов обработки SDS и Triton X-100 культуры с ранней логарифмической фазой инокулировали (1%, об. / Об.) В бульон MRS, содержащий 0,01% или 0,02% (мас. / Об.) SDS или 0,25% или 0,50% (об. / Об.). т.) Тритон Х-100. Рост оценивали путем определения OD 600 после 24 ч инкубации при 37 ° C. Для испытания замораживанием-оттаиванием порции по 1 мл культур с ранней логарифмической фазой центрифугировали и ресуспендировали в равном объеме 0,1 × бульона MRS. Образцы подвергали многократным циклам замораживания (баня этанол-сухой лед, 3 мин) и оттаивания (водяная баня 37 ° C, 3 мин).Выживших оценивали путем посева на агар MRS после 5, 10, 15 и 20 циклов замораживания-оттаивания.
Анализ адгезии эпителиальных клеток Caco-2. Для анализа адгезии клеток Caco-2 клеточную линию Caco-2 (ATCC HTB-37) получали из Американской коллекции типовых культур (Роквилл, Мэриленд). Все среды для культивирования клеток и реагенты были приобретены у Gibco (Gibco-Invitrogen Corp., Карлсбад, Калифорния). Клетки Caco-2 получали и поддерживали для исследований адгезии, как описано ранее Buck et al. (20) со следующими модификациями.Вкратце, клетки обычно выращивали при 37 ° C в атмосфере 95% воздуха и 5% CO 2 в минимальной необходимой среде с добавлением 1 мМ пирувата натрия, 20% (об. / Об.) Инактивированного нагреванием (56 ° C, 30%). мин) фетальная бычья сыворотка, 0,1 мМ заменимых аминокислот и антибиотики (100 мг / мл пенициллина G, 100 мг / мл сульфата стрептомицина и 0,25 мг / мл амфотерицина). Для анализа адгезии монослои готовили в 12-луночных планшетах для культивирования тканей путем засева приблизительно 6,5 × 10 4 клеток на лунку 2 мл среды для культивирования клеток.Культуральную среду заменяли каждые 2 дня, и монослои использовали для анализа приверженности через 14-15 дней после контакта. Клетки использовали между пассажами 23 и 40. Перед анализами на адгезию монослои дважды промывали 1 мл фосфатно-солевого буфера (PBS) (pH 7,4; Invitrogen), а затем к каждому добавляли 1 мл свежей культуральной среды без антибиотиков. хорошо.
Для приготовления бактериальных клеток 10 мл клеток средней логарифмической фазы (OD 600 , 0,6–0,8), выращенных в бульоне MRS, осаждали центрифугированием при 3,166 × г в течение 10 мин при комнатной температуре, промывали один раз. с 10 мл PBS и ресуспендировали в PBS до конечной концентрации ~ 4 × 10 8 КОЕ / мл.Один миллилитр бактериальной суспензии добавляли в каждую лунку, содержащую монослой Caco-2, и инкубировали при 37 ° C в течение 1 часа. После этой инкубации монослои промывали пять раз 1 мл PBS для удаления несвязавшихся бактерий, а затем инкубировали с 1 мл 0,05% (об. / Об.) Triton X-100 в течение 10 мин. Суспензии однослойных клеток и бактерий серийно разводили в 0,1 × бульоне MRS и высевали на агар MRS для определения количества прикрепившихся клеток (выраженных в КОЕ). Все эксперименты проводились по крайней мере с тремя независимыми культурами, каждая с трехкратной повторностью лунок, содержащих монослои.
Анализ адгезии муцина. Для иммобилизации муцина желудка свиньи (тип III; Sigma) на 96-луночном круглодонном планшете из полистирола Costar (Corning Inc., Нью-Йорк) использовали 10 мг / мл раствора муцина в дистиллированной воде. готовили, разливали в каждую лунку (100 мкл на лунку) и инкубировали при 4 ° C в течение ночи. Избыток муцина удаляли двумя промывками 200 мкл PBS перед добавлением бактериальных клеток. Культуры в средней логарифмической фазе, выращенные в бульоне MRS, один раз промывали PBS, и конечную OD 600 доводили до 0.6 с PBS. Клеточную суспензию (100 мкл) добавляли в каждую лунку и инкубировали при 37 ° C в течение 1 часа. Затем каждую лунку пять раз промывали 200 мкл PBS для удаления несвязавшихся клеток, после чего следовала обработка 200 мкл 0,05% Triton X-100 для отделения слоя муцина и бактериальных клеток. Суспензию клеток разводили в 0,1 × бульоне MRS и высевали на агар MRS для определения количества прилипших клеток (выраженного в КОЕ). Все эксперименты проводились с двумя независимыми культурами, каждая из которых имела трехкратные лунки, содержащие слой муцина.
Анализ микробной адгезии к растворителям (MATS). Анализ микробной адгезии к растворителям (MATS) проводили на основе протокола, описанного Bellon-Fontaine et al. (12) со следующими двумя парами растворителей: (i) хлороформ (кислота Льюиса, акцептор электронов, полярный) и гексадекан (аполярный), и (ii) этилацетат (основание Льюиса, донор электронов, полярный) и декан (неполярный). ). Вкратце, 5-миллилитровые порции культур средней логарифмической фазы, выращенных в бульоне MRS, собирали центрифугированием при 3166 × г при комнатной температуре, промывали один раз в равном объеме PBS и ресуспендировали в PBS до конечной OD 400. из 0.8. Затем 1,2 мл каждой клеточной суспензии смешивали с 0,2 мл каждого растворителя путем встряхивания в течение 60 с. Смеси инкубировали при комнатной температуре в течение 15 мин, чтобы обеспечить полное разделение двух фаз. Затем 600 мкл водной фазы осторожно удаляли для определения OD 400 . Уровень сродства клеток к растворителям рассчитывали по следующему уравнению:% аффинности = 100 × [1 — ( A / A 0 )], где A 0 и A являются OD 400 суспензии клеток до и после смешивания с растворителями, соответственно.Все анализы были выполнены в двух экземплярах с двумя независимыми культурами.
Сканирующая электронная микроскопия (SEM). Клетки выращивали в среде MRS (14 мл) до OD 600 от 0,6 до 0,8 (при этом pH супернатанта культуры составлял от ~ 5,1 до 5,3) и осаждали центрифугированием при 3166 × . g при комнатной температуре. Осадки клеток ресуспендировали в свежей фиксирующей смеси 1: 1 (об. / Об.), Содержащей 6% глутарового альдегида и 0,2 М какодилата натрия (pH 5,5), и хранили при 4 ° C. Обработка образцов для сканирующей электронной микроскопии (SEM) была выполнена Центром электронной микроскопии при Университете штата Северная Каролина.Образцы просматривали с помощью растрового электронного микроскопа JOEL JSM 5900LV при 15 кВ.
Номера доступа к данным микрочипов. Платформа микрочипов и данные серий доступны на сайте Gene Expression Omnibus (http://www.ncbi.nlm.nih.gov/geo ) под инвентарными номерами GPL7426 (платформа) и GSE14622 (серия).
РЕЗУЛЬТАТЫ
Конструирование мутанта NCFM Δ upp в качестве хозяина для контрселективной аллельной замены. Чтобы исследовать потенциал контрселективной стратегии на основе upp для L.acidophilus NCFM, этот штамм впервые был протестирован для определения его чувствительности к 5-ФУ. Ночные культуры в бульоне MRS серийно разводили в SDM и высевали при плотности ок. 10 5 КОЕ на чашку на SDM, содержащем различные концентрации 5-ФУ или эквивалентные количества ДМСО (агента, используемого для растворения 5-ФУ) в качестве контроля. Рост подавлялся на среде, содержащей 100 мкг / мл 5-ФУ. Присутствие ДМСО в среде не влияло на рост L. acidophilus , указывая на то, что ингибирование роста было обусловлено исключительно токсичностью 5-ФУ.Следовательно, 100 мкг / мл 5-ФУ было приемлемой концентрацией для отбора против интегрантов, которые не разрешили меродиплоидное состояние.
Чувствительность L. acidophilus к 5-ФУ подтвердила присутствие функциональной UPRTase. Предполагаемый ген, кодирующий UPRTase, LBA0770, был обнаружен в последовательности генома NCFM. Кодирующая область LBA0770 была аннотирована как upp на основании сходства последовательностей с соответствующими ферментами в базе данных GenBank. Выведенная аминокислотная последовательность показала более 80% идентичности последовательности с UPRTases из L.delbrueckii, L. gasseri и L. johnsonii . Чтобы связать генотип upp с наблюдаемым фенотипом 5-FU s , с использованием двухэтапной схемы замещения аллелей на основе pORI была сконструирована делеция 315 п.н. в рамке считывания в предполагаемом гене upp длиной 630 п.н. (см. Материалы и методы). После размножения хромосомного интегранта L. acidophilus pTRK934 в неселективной среде для облегчения второй гомологичной рекомбинации 1200 колоний были проверены на рекомбинанты Em s , которые потеряли остов плазмиды.Из 13 выделенных колоний Em s , 4 несли удаленный аллель Δ upp , что подтверждено анализом ПЦР и секвенированием ДНК целевого локуса. Все четыре мутанта Δ upp были способны расти на SDM в присутствии 100 мкг / мл 5-ФУ, концентрация, которая подавляет рост дикого типа. Этот результат подтвердил, что LBA0770 кодирует функциональную UPRTase в NCFM. Все мутанты Δ upp не показали видимых различий в скорости роста по сравнению с родительским штаммом NCFM, когда они культивировались в бульоне MRS (данные не показаны), что указывает на то, что делеция в локусе upp не влияла на рост. поведение мутантов Δ upp .Один из мутантов Δ upp , обозначенный NCK1909, был выбран в качестве фонового хозяина для контрселективной замены гена.
PM анализ мутанта NCK1909 upp Δ . Поскольку мутант NCK1909 служит как хозяином для генетического обмена, так и эталонным штаммом для последующих фенотипических исследований делеционных мутантов, которые будут получены с помощью системы контрселективного нокаута гена, мы использовали Biolog PM для обнаружения любых фенотипических различий между мутантом Δ upp и его родительским штаммом NCFM.Этот анализ позволил нам сравнить фенотипы двух штаммов в ответ на осмолиты, pH и 240 различных химических субстратов. В этом анализе только 1 из 1152 скважин показала разницу в росте (PM19, D07) (см. Рис. S1 и Таблицу S3 в дополнительном материале). Однако разница, наблюдаемая для роста в присутствии йодонитротетразолия фиолетового, не считалась значимой, поскольку разница в росте наблюдалась только при одной из четырех испытанных концентраций. Никаких различий не наблюдалось для pH, осмолитов или другой химической чувствительности.За исключением устойчивости к 5-ФУ, результаты продемонстрировали, что мутация upp не вызвала каких-либо серьезных обнаруживаемых различий в клеточном фенотипе по сравнению с родительским штаммом.
Исследование экспрессии генов на микрочипах у фонового хозяина NCK1909. Ранее мы успешно разработали микроматрицу для полногенома L. acidophilus NCFM, покрывающую примерно 97,4% открытых рамок считывания, предсказанных в геноме (7). В настоящем исследовании мы разработали новую платформу полногеномного oligoarray NCFM с использованием OligoArray 2.1 (45). В общей сложности 1823 олигонуклеотида (представляющих 97,8% генов в хромосоме), каждый из которых представляет собой ген, наносили шесть раз случайным образом на предметные стекла (общее количество пятен 11520). Чтобы оценить надежность платформы, мы провели эксперимент по гибридизации микрочипов, пометив кДНК из того же образца РНК с помощью Cy3 и Cy5. Данные гибридизации выявили линейную корреляцию (значение R ) в относительных уровнях экспрессии 0.98 для 11520 пятен со средним, максимальным и минимальным отношениями экспрессии Cy5 / Cy3 1,0, 1,48 и 0,46, соответственно (данные не показаны).
Для дальнейшего установления генотипа NCK1909 в качестве репрезентативного эталонного штамма была проведена гибридизация микрочипов с использованием новой платформы oligoarray для сравнения профилей транскрипции NCK1909 с профилями NCFM в раннем журнале (OD 600 , 0,3) и в середине. log (OD 600 , 0,8), когда организмы культивировали в среде MRS при 37 ° C в условиях окружающей атмосферы.Эти плотности клеток представляют собой стадии роста, когда клетки собирали для анализов стрессового заражения (OD 600 , от 0,25 до 0,3) и анализов на адгезию клеток (OD 600 , от 0,6 до 0,8) (см. Материалы и методы). Анализ данных экспрессии генов обнаружил только несколько генов, которые по-разному экспрессировались посредством однократного или большего изменения NCK1909 и NCFM как на ранней, так и на средней логарифмической фазах, когда организмы выращивались в среде MRS (таблица 2). Кроме того, не было обнаружено ни одного гена, экспрессирующегося в NCK1909 на уровне, более чем в один раз превышающем уровень NCFM во время фазы роста в середине логарифма.В целом, эти результаты дополнительно подтверждают, что NCK1909 является репрезентативным эталонным штаммом, который можно использовать как для фенотипических, так и для генотипических исследований делеционных мутантов, полученных с использованием системы контрселективной замены гена на основе upp .
ТАБЛИЦА 2.Сравнительные профили экспрессии генов NCFM и Δ upp мутанта NCK1909, выращенного в среде MRS
Конструирование вектора интеграции на основе pORI. Вектор pORI28 был выбран в качестве основы для создания вектора интеграции с возможностью обратного отбора. из-за его установленной эффективности для создания хромосомных вставок и делеций в NCFM.Предыдущий анализ транскриптома NCFM, выращенного на различных источниках углеводов, показал, что ген upp был высоко экспрессирован во всех экспериментальных условиях (10). Следовательно, нативный промотор upp использовали для конститутивной экспрессии эктопического гена upp . Ген lacZ ‘из pUC19 также был включен в вектор интеграции, который содержит обычную MCS и обеспечивает скрининг сине-белого цвета для облегчения отбора рекомбинантов E. coli .
Чтобы сконструировать вектор контрселективной интеграции, всю область множественного клонирования pORI28 удалили рестрикцией BglII и XbaI, затупили концы и лигировали с продуктами SOE-PCR P- upp и lacZ ‘гена с его вышележащий промотор и сайт связывания CAP амплифицированы из вектора pUC19 (рис. 1). Секвенирование полученного вектора интеграции, pTRK935, неожиданно показало присутствие интактного сайта XbaI и пяти дополнительных случайных оснований в месте лигирования между lacZ ‘и остовом pORI28, предположительно в результате обработки тупым концом расщепленного pORI28.Таким образом, сайт рестрикции XbaI в области множественного клонирования pTRK935 не функционирует как уникальный сайт для клонирования гена. Когда pTRK935 вводили в NCK1909, несущий вспомогательную плазмиду pTRK669 (NCK1910), не наблюдалось роста в присутствии 100 мкг / мл 5-FU. Этот результат подтвердил, что P- upp в плазмиде способен дополнять генотип Δ upp в хозяине интеграции NCK1910 и восстанавливать чувствительность к 5-FU и что он может служить функциональным маркером для поддержания отрицательной селекции против меродиплоидов. с 5-ФУ.
Анализ последовательности гена slpX . Для оценки эффективности стратегии контрселекции на основе upp был выбран ген slpX для нацеленной делеции. L. acidophilus NCFM имеет два основных гена белка S-слоя: slpA (LBA0169) и молчащий ген slpB (LBA0175) (таблица 3) (32). Наше недавнее профилирование NCFM S-слоя на основе MS-анализа пептидов, полученных из очищенных S-слоев, выявило коэкспрессию белка SlpX, кодируемого slpX , с SlpA (экспрессируемым штаммом дикого типа) и SlpB (экспрессируемым с помощью slpA ). Мутант со вставкой , NCK1377) (см. Таблицу S4 в дополнительном материале).Таким образом, предполагается, что SlpX вносит вклад в комплекс S-слоя в NCFM. Гены slpA и slpB расположены в кластере генов, тогда как slpX находится в удаленном хромосомном локусе. Выведенная последовательность SlpX имела умеренную идентичность последовательности (53%) с последовательностью белка поверхностного слоя из L. acidophilus JCM1038, единственного ортолога SlpX, найденного в базе данных GenBank. Между тем, SlpX имел менее 30% идентичности последовательности с SlpA (26%) и SlpB (24%), и сходство ограничивалось N-концевой и C-концевой областями.Тем не менее, SlpX похож по размеру на SlpA и SlpB (от 46 до 51 кДа), и все три белка S-слоя имеют общие характеристики, такие как предсказанная базовая изоэлектрическая точка ∼9,5 и высокая доля гидрофобных остатков (от 44 до 48%). ) (Таблица 3), которые являются типичными характеристиками белков S-слоя лактобацилл.
ТАБЛИЦА 3.Общая характеристика белков S-слоя в L. acidophilus NCFM
Делеция в рамке считывания гена slpX с использованием безмаркерной замены гена контрселекцией на основе upp .Для конструирования делеции в рамке считывания 1356 п.н. (~ 90% гена) в гене slpX , примерно 750 п.н. хромосомные сегменты, фланкирующие обе стороны области-мишени делеции, амплифицировали с помощью ПЦР, соединенных с помощью SOE-PCR. и клонировали в сайты BamHI / SacI pTRK935. Полученная конструкция, pTRK956, была электропорирована в L. acidophilus NCK1910, и трансформанты были отобраны на основе устойчивости к Em и Cm (фиг. 2). Один трансформант pTRK956 был отобран и размножен в бульоне MRS при 42 ° C в присутствии Em в течение 30 поколений для облегчения интеграции pTRK956 в хромосому посредством гомологичной рекомбинации.Затем интегрированную плазмиду отделяли от хромосомы посредством вторичной рекомбинации, пропуская интегрант в бульоне MRS без Em при 37 ° C в течение 50 поколений. Рекомбинанты, не содержащие плазмид, выделяли на SDM с 5-FU с частотой приблизительно 1,4 × 10 -3 . Девять из 18 выбранных изолятов 5-FU r несли удаленный аллель Δ slpX , тогда как остальные изоляты сохранили аллель дикого типа (рис. 3). Все рекомбинанты Δ slpX были Em s , что подтверждает удаление интегрированного остова pTRK956 из хромосомы.Генотип подтверждали секвенированием ДНК. Один из мутантов Δ slpX , обозначенный NCK1962, был выбран для дальнейшего изучения.
РИС. 2. Стратегия безмаркерной замены гена с upp в качестве контрселективного маркера для делеции в рамке считывания 1356 п.н. в гене slpX . Рекомбинация pTRK956 с одним кроссовером произошла в хромосомной области slpX , гомологичной фрагменту A или B (показана только рекомбинация через фрагмент A). Удаление селекции Em для интегрированной плазмиды pTRK956 облегчает вторую рекомбинацию и удаление плазмиды, генерируя рекомбинанты либо с аллелем дикого типа, либо с аллелем Δ slpX .Полученные рекомбинанты могут быть отобраны на SDM, содержащем 100 мкг / мл 5-FU. Фрагменты ДНК A (пунктирный прямоугольник) и B (заштрихованный прямоугольник) представляют клонированную копию аллеля Δ slpX , где фрагмент A состоит из 5′-области slpX (105 п.н.) и расположенной выше фланкирующей области и фрагмента B. состоит из 3′-области slpX (36 п.н.) и нижележащей фланкирующей области. РИС. 3.ПЦР-скрининг генотипа Δ slpX в случайных колониях, выделенных после отбора 5-ФУ.(A) Генная организация области, окружающей локус slpX . Стрелки указывают расположение праймеров slpX-up и slpX-dw, используемых для анализа ПЦР. Заштрихованная область в гене slpX представляет собой мишень для делеции. Шпильчатые структуры указывают на предполагаемые rho-независимые терминаторы. Хромосомная карта нарисована не в масштабе. (B) ПЦР-анализ колоний 18 отобранных рекомбинантов 5-FU r . Ожидаемые размеры ампликонов, генерируемых из генотипов дикого типа и Δ slpX , составляют приблизительно 3.14 кб и 1,78 кб соответственно. Дорожка М, маркер размера ДНК; дорожки с 1 по 18, изоляты 5-FU r ; дорожка WT, родительский штамм NCFM (контроль).
Профиль S-слоя мутанта NCK1962. Разложение очищенных белков S-слоя с помощью электрофореза в SDS-полиакриламидном геле (PAGE) подтвердило отсутствие полосы 51 кДа, соответствующей белку SlpX в NCK1962 (рис. 4). Как и ожидалось, мутант экспрессировал только доминантные белки SlpA (46 кДа), хотя и на несколько более высоком уровне, чем штамм дикого типа, исходя из относительной интенсивности полос.Кроме того, не было обнаружено SlpX-специфического пептида для NCK1962 на основании исследований MS пептидов, полученных в результате трипсинолиза очищенных S-слоев (см. Таблицу S4 в дополнительном материале). Эти результаты дополнительно подтвердили, что отсутствие белка SlpX в S-слоях NCK1962 было связано с делецией гена slpX . Между тем, как для штамма NCFM, так и для интегранта NCK1377 общая интенсивность полос экспрессируемых белков S-слоя свидетельствует о том, что SlpX присутствует на значительно более низком уровне в комплексе S-слоя, чем белки SlpA или SlpB.
РИС. 4.SDS-PAGE очищенных белков S-слоя из штамма NCFM дикого типа L. acidophilus (дорожка WT), интегранта NCFM slpA NCK1377 (дорожка 1377) и мутанта с делецией slpX NCK1962 (дорожка 1962), использованный для анализа МС. Профиль S-слоя каждого штамма суммирован внизу на основе результатов МС-анализа очищенных белков S-слоя. +, присутствует; -, отсутствует. Слева указаны молекулярные массы стандартных белков (дорожка M).
Фенотип мутанта NCK1962.Чтобы исследовать роль SlpX в ассоциации с S-слоем у L. acidophilus , мы исследовали влияние делеции slpX на морфотип клеток, реакцию на стрессы окружающей среды и свойства клеточной поверхности. Мониторинг роста в среде MRS, основанный на измерении плотности клеток и количества клеток, выявил несколько более медленный рост мутанта NCK1962, чем эталонного штамма NCK1909, а максимальные удельные скорости роста составили 0,43 часа -1 (после 9 часов роста) и 0.56 ч −1 (через 7 ч роста) соответственно (рис. 5). Тем не менее, культуры NCK1962 в конечном итоге достигли конечной плотности клеток, аналогичной плотности культур эталонного штамма. Исследования NCK1962 под микроскопом и с помощью SEM не показали значительных различий в морфологии и внешнем виде клеток по сравнению с NCK1909 (данные не показаны). На основании анализа MATS оба штамма показали высокое сродство (от 80 до 95%) к неполярным растворителям гексадекану и декану (данные не показаны), что указывает на то, что мутант NCK1962 сохранил гидрофобную клеточную поверхность.Кроме того, NCK1962 проявлял основные свойства клеточной поверхности, аналогичные свойствам NCK1909, о чем свидетельствует его более высокое сродство к хлороформу (> 99%), чем к этилацетату (30%).
РИС. 5.Рост L. acidophilus NCK1962 и NCK1909 в среде MRS при 37 ° C в условиях окружающей атмосферы. Рост оценивали путем определения оптической плотности и количества клеток (КОЕ / мл).
Когда культуры контрольного штамма NCK1909 и мутантного штамма NCK1962 подвергались воздействию 0.3% свиной желчи в течение 2 часов, мутант имел более высокую выживаемость (23,5% ± 3,9%), чем NCK1909 (2,6% ± 0,8%) (фиг. 6A). Точно так же NCK1962 был более устойчив к обработке 2,5% бычьей желчью; 57,1% ± 12,9% выживших выздоровели, по сравнению с только 4,3% ± 0,7% жизнеспособных клеток NCK1909. С другой стороны, NCK1962 был более восприимчив к SDS, и сильное ингибирование роста наблюдалось при 0,02% SDS (фиг. 6B). Между тем, NCK1962 показал рост, сравнимый с ростом NCK1909 в присутствии Triton X-100.Кроме того, не наблюдалось значительной разницы в выживаемости NCK1962 и NCK1909 при заражении организмов 10% NaCl, 15% этанолом, низким pH (pH 3,0 или 3,5) в присутствии органической кислоты (молочной кислоты) или неорганическая кислота (HCl) или повторяющиеся циклы замораживания-оттаивания (данные не показаны). Два штамма также продемонстрировали сопоставимые способности прикрепляться к эпителиальным клеткам Caco-2 и муцину желудка свиньи in vitro (данные не показаны).
РИС. 6.Выживание NCK1962 (мутант Δ slpX ) и NCK1909 (эталонный штамм) после анализов с нагрузкой на стресс.(A) Выживание культур NCK1909 и NCK1962 в ранней логарифмической фазе после воздействия среды MRS с добавлением 0,3% желчи свиньи или 2,5% желчи быка в течение 2 часов при 37 ° C в условиях окружающей атмосферы. (B) Культуры NCK1909 и NCK1962 в ранней логарифмической фазе инокулировали (1% инокулята) в бульон MRS с добавлением SDS или поверхностно-активного вещества Triton X-100, и плотность клеток измеряли через 24 часа инкубации при 37 ° C в условиях окружающей атмосферы. условия. Данные представляют собой средние значения ± стандартная ошибка средних для трех независимых повторов.
ОБСУЖДЕНИЕ
L. acidophilus принадлежит к одной из самых динамичных групп комменсалов среди обширного консорциума микрофлоры, населяющей верхние отделы желудочно-кишечного тракта человека. Сравнительный геномный анализ с другими видами Lactobacillus и исследования транскрипции микрочипов выявили важные особенности генов, которые могут способствовать выживанию, адаптации и колонизации L. acidophilus в сложной кишечной среде (2, 6, 7, 10, 21). , 31, 42).В связи с промышленным значением L. acidophilus в качестве пробиотика и диетического дополнения, обширные исследования также были посвящены прогнозированию ключевых генов, которые играют важную роль в выживании и активности этого организма как в биопроцессорной среде, так и в среде хозяина. Постгеномный анализ L. acidophilus с целью установления функциональных ролей этих генов и связанных регуляторных сетей требует эффективных инструментов генетического обмена, поскольку чаще всего создание нокаутов генов после идентификации представляющих интерес генных мишеней представляет собой основное узкое место. в функциональных геномных исследованиях.
В этом отчете разработка стратегии встречного отбора на основе upp описана для NCFM L. acidophilus с целью повышения эффективности традиционной системы нокаута гена pORI путем обеспечения прямого отбора для свободных от плазмид рекомбинантов с использованием 5-FU. . Ген upp , кодирующий UPRTase (EC 2.4.2.9), участвует в пути восстановления пиримидина, поскольку UPRTase превращает урацил в UMP для биосинтеза нуклеотидов. Удаление upp в NCFM придает устойчивость к 5-FU, подтверждая присутствие функциональной UPRTase.Цитотоксичность 5-FU инициируется превращением 5-FU в 5-фтор-UMP с помощью UPRTase, что приводит к последующему образованию очень токсичного метаболита 5-фтордезоксиуридинмонофосфата (5-FdUMP) (40). Последнее соединение действует как ингибитор тимидилатсинтазы и тем самым предотвращает образование dTMP, необходимого для синтеза предшественника dTTP. У некоторых микроорганизмов 5-FU также может напрямую превращаться в 5-FdUMP тимидинфосфорилазой и тимидинкиназой (3, 36, 37). Предыдущий анализ генома показал, что L.acidophilus NCFM является ауксотрофным по UTP и dTTP (2). Отсутствие генов, кодирующих нуклеозиддифосфаткиназу и нуклеозидтрифосфатаденилаткиназу, необходимых для превращения UDP (из UMP) в UTP, а также тимидинфосфорилазу, может предотвратить образование 5-FdUMP. Поэтому мы предположили, что фенотип NCFM 5-FU s , вероятно, обусловлен кумулятивным токсическим действием промежуточных продуктов 5-фтор-UMP или 5-фтор-UDP. Это наблюдение также подразумевает, что, поскольку NCFM не имеет полного пути биосинтеза пиримидина de novo, мутация upp не приведет к значительным различиям в физиологическом поведении и метаболизме урацила между двумя штаммами.Эта гипотеза подтверждается фенотипическим анализом микроматрицы, сравнивающим исходный штамм NCFM и фоновый хозяин Δ upp NCK1909, где не наблюдали фенотипических различий в ответ на pH, осмолиты и исследуемые химические субстраты. Кроме того, микроматричный анализ транскрипции NCFM и NCK1909, выращенных в среде MRS, показал небольшую разницу в глобальных профилях экспрессии генов между этими двумя штаммами. В совокупности эти результаты подтверждают использование хозяина NCK1909 в качестве представителя штамма NCFM дикого типа для фенотипических исследований in vitro и in vivo мутантов, сконструированных с этой системой замены генов.
Для оценки эффективности контрселективного подхода upp в отношении L. acidophilus была использована стратегия нокаута для создания делеции в рамке считывания в гене slpX . SlpX представляет собой секретируемый белок, который недавно был идентифицирован как белок, связанный с комплексом S-слоя в L. acidophilus NCFM. В общем, S-слои представляют собой паракристаллические слои (глико) белковых субъединиц, которые в изобилии присутствуют на поверхности клеток большинства эубактерий и архей (для обзора см. Ссылку 48).S-слои были идентифицированы и охарактеризованы у множества видов Lactobacillus (для обзора см. Ссылку 4). Примечательно, что L. brevis и несколько видов, тесно связанных с L. acidophilus , включая L. crispatus, L. amylovorus и L. gallinarum , относятся к лактобактериям, продуцирующим S-слой, которые имеют несколько S гены, кодирующие белок (16-18, 29). Тем не менее, только белок поверхностного слоя из L. acidophilus JCM1038, который имел умеренное сходство последовательностей с SlpX, был обнаружен в текущей базе данных GenBank (см. Выше).Возникает соблазн предположить, что белок SlpX является уникальным для NCFM, хотя еще предстоит определить, присутствует ли гомолог SlpX у несеквенированных видов, которые имеют S-слои. Для S-слоев были предложены различные роли, в том числе роли детерминант формы клеток, молекулярных сит, слоев, защищающих от адсорбции фагов или хищных бактерий, и слоев, обеспечивающих места закрепления для связанных с поверхностью ферментов, адгезии клеток и распознавания поверхности. Предполагается, что большинство этих функций важны для выживания L.acidophilus в желудочно-кишечном тракте человека и для клеточной коммуникации с иммунными клетками хозяина и эпителиальными клетками слизистых оболочек. Из-за функционального потенциала S-слоев и тесной ассоциации SlpX с SlpA или SlpB, было очень интересно исследовать функциональную роль SlpX в образовании комплекса S-слоев.
Мутация гена slpX не повлияла на морфологию мутанта NCK1962, хотя мутантный штамм показал немного более низкую скорость роста, чем фоновый хозяин NCK1909.Предыдущее исследование van der Mei et al. (50) продемонстрировали, что S-слои придают лактобацилл гидрофобность клеточной поверхности. В настоящем исследовании отсутствие белка SlpX в структуре S-слоя NCK1962 не вызывало значительного изменения гидрофобности или основных свойств клеточной поверхности на основе ее сродства к растворителям. Одно из возможных объяснений состоит в том, что белки SlpX в природе присутствуют на гораздо более низких уровнях, чем SlpA или SlpB, как показал анализ очищенных S-слоев родительского штамма и SlpB-экспрессирующего интегранта slpA NCK1377 (рис.4). Таким образом, отсутствие SlpX в NCK1962 может иметь незначительное влияние на общие гидрофобные и основные свойства клеточной поверхности. Альтернативно, изменение свойств поверхности из-за отсутствия SlpX может быть компенсировано сверхэкспрессией SlpA в мутанте NCK1962.
Никаких серьезных различий между NCK1962 и NCK1909 не наблюдалось, когда они подвергались ряду стрессовых воздействий окружающей среды, таких как присутствие 10% NaCl или 15% этанола или низкий pH.NCK1962 также показал выживаемость, аналогичную NCK1909, после повторной обработки свободным оттаиванием. Эти результаты предполагают, что отсутствие SlpX в комплексе S-слоя не ставит под угрозу способность мутанта справляться с осмотическим стрессом и стрессом этанола или поддерживать гомеостаз pH или физическую целостность после повреждения замораживанием-оттаиванием. Бак и др. (20) недавно показали, что мутант slpA имел значительно сниженную способность прикрепляться к эпителиальным клеткам Caco-2 in vitro. Эти авторы, однако, указали, что наблюдаемый фенотип пониженной адгезии у мутанта slpA также может быть следствием отсутствия поверхностных белков, ассоциированных с S-слоем, которые вносят вклад в адгезию.В настоящем исследовании мутант NCK1962 не показал сниженной способности прикрепляться к эпителиальным клеткам Caco-2 или муцину in vitro. Это означает, что SlpX не принимает непосредственного участия в поверхностной адгезии или что он может играть второстепенную роль как место закрепления для потенциальных факторов адгезии.
Неожиданно отсутствие белков SlpX в комплексе S-слоя мутанта NCK1962 сделало клетки более восприимчивыми к анионному поверхностно-активному веществу SDS, но не к неионогенному поверхностно-активному веществу Triton X-100, чем эталонный штамм.Обычно оба поверхностно-активных вещества оказывают бактерицидное действие, вызывая лизис клеток. Из-за анионных свойств SDS, которые разрушают нековалентные связи в белках, наблюдаемый SDS-чувствительный фенотип NCK1962 мог быть частично обусловлен денатурацией, вызванной SDS, основных поверхностных белков или транспортеров в NCK1962, которые в противном случае были защищены интактным S-образцом. слой с SlpX в NCK1909. С другой стороны, NCK1962 продемонстрировал значительно повышенную толерантность как к биологическим поверхностно-активным веществам, так и к бычьей желчи (бычьей желчи) и свиной желчи, тогда как NCK1909 наблюдалось почти 2-логарифмическое снижение при тех же обработках.Основными органическими составляющими желчи являются соли желчных кислот, холестерин и фосфолипиды (обзор см. В ссылке 11). Соли желчных кислот синтезируются de novo печенью из холестерина, где стероидная часть молекулы конъюгирована либо с глицином (гликоконъюгированным), либо с таурином (тауроконъюгированным) через пептидные связи. Мы предположили, что отсутствие SlpX может повлиять на проницаемость S-слоев. Это потенциально позволило бы увеличить внутриклеточный доступ конъюгированных солей желчных кислот. Возможно, ожидаемое изменение проницаемости SlpX-дефицитных S-слоев в NCK1962 может потенциально позволить более эффективный отток внутриклеточных деконъюгированных желчных кислот.
На данный момент точная роль slpX остается неясной. Более низкое содержание SlpX в S-слоях, выявленное с помощью SDS-PAGE, позволяет предположить, что этот белок является вспомогательным компонентом комплекса S-слоя. Тем не менее, мы пришли к выводу, что SlpX важен для поддержания клеточной целостности и других неизвестных физиологических функций. Это связано с наблюдениями, что инактивация этого белка влияет на скорость роста и толерантность к солям желчных кислот в мутанте NCK1962, а также на его относительную сверхэкспрессию в NCK1377, SlpB-доминантном штамме, в котором отсутствует белок SlpA.Это означает, что SlpX способствует выживанию и пробиотической функциональности в среде GI хозяина. Дальнейшие фенотипические исследования и исследования экспрессии генов в настоящее время проводятся для установления функции SlpX. В совокупности результаты настоящего исследования проливают свет на сложность биологических ролей S-слоев у L. acidophilus .
В целом, мы продемонстрировали функциональность стратегии контрселекции upp для безмаркерного аллельного обмена в L.acidophilus путем конструирования делеции в рамке считывания в гене slpX . На сегодняшний день эта система использовалась в нашей лаборатории для создания множественных делеций генов, нацеленных на один или несколько локусов гена в L. acidophilus NCFM. Стратегия встречного отбора позволяет прямой отбор двойных рекомбинантов, тем самым облегчая конструирование мутантов и назначение ролей генам-кандидатам, которые важны для пробиотических свойств L. acidophilus .Поскольку контрселективный маркер и upp пригоден для вторичной переработки, мутации в бесчисленных локусах генов могут быть последовательно введены в один и тот же штамм для изучения сложных регуляторных сетей генов. Кроме того, мутации могут быть нацелены на гены оперона без полярного воздействия на соседние гены. Эта улучшенная контрселективная система замены аллелей также должна оказаться полезной для введения желаемых пробиотических признаков в L. acidophilus или для создания новых штаммов для доставки биотерапевтических средств.
БЛАГОДАРНОСТИ
Эта работа была частично поддержана Danisco USA, Inc. (Мэдисон, Висконсин), North Carolina Dairy Foundation и Dairy Management, Inc.
Мы очень благодарны Кристофу Брандили за его техническую помощь , Сабрине Тачджиан и Р. Келли за то, что они поделились своим протоколом гибридизации олигомассивов, и Розмари Санозски-Дауэс и Ричардом Таллон за их помощь и обсуждения, касающиеся эпителиальных клеток Caco-2 in vitro, исследований адгезии муцина и анализа MATS.
- Copyright © 2009 Американское общество микробиологов.
СПРАВОЧНАЯ ИНФОРМАЦИЯ
- 1.10
Альтерманн, Э., Б. Л. Бак, Р. Кано и Т. Р. Клаенхаммер. 2004. Идентификация и фенотипическая характеристика белка клеточного деления CdpA. Gene342 : 189-197.
- 2.↵
Альтерманн, Э., У. М. Рассел, М. А. Азкарат-Перил, Р. Баррангу, Б. Л. Бак, О. Маколифф, Н. Саутер, А. Добсон, Т. Дуонг, М.Калланан, С. Лик, А. Хэмрик, Р. Кано и Т. Р. Клаенхаммер. 2005. Полная последовательность генома пробиотической молочнокислой бактерии Lactobacillus acidophilus NCFM. Proc. Natl. Акад. Sci. USA102 : 3906-3912.
- 3.↵
Arsene-Ploetze, F., H. Nicoloff, B. Kammerer, J. Martinussen и F. Bringel. 2006. Путь спасения урацила в Lactobacillus plantarum : транскрипция и генетические исследования. J. Bacteriol. 188 : 4777-4786.
- 4.↵
Avall-Jaaskelainen, S., and A. Palva. 2005. Поверхностные слои Lactobacillus и их применение. FEMS Microbiol. Ред.29 : 511-529.
- 5.↵
Азкарат-Перил, М. А., Э. Альтерманн, Р. Л. Гувер-Фитцула, Р. Кано и Т. Р. Клаенхаммер. 2004. Идентификация и инактивация генетических локусов, связанных с устойчивостью к кислоте Lactobacillus acidophilus . Прил. Environ.Microbiol.70 : 5315-5322.
- 6.↵
Азкарат-Перил, М. А., Дж. М. Бруно-Барсена, Х. М. Хассан и Т. Р. Кленхаммер. 2006. Транскрипционный и функциональный анализ генов оксалил-кофермента А (КоА) декарбоксилазы и формил-КоА трансферазы из Lactobacillus acidophilus . Прил. Environ. Microbiol.72 : 1891-1899.
- 7.↵
Azcarate-Peril, M. A., O. McAuliffe, E. Altermann, S.Лик, В. М. Рассел и Т. Р. Кленхаммер. 2005. Анализ микроматрицы двухкомпонентной регуляторной системы, участвующей в кислотной устойчивости и протеолитической активности у Lactobacillus acidophilus . Прил. Environ. Microbiol.71 : 5794-5804.
- 8.↵
Barefoot, S. F., and T. R. Klaenhammer. 1983. Обнаружение и активность лактацина B, бактериоцина, продуцируемого Lactobacillus acidophilus . Прил. Environ. Microbiol.45 : 1808-1815.
- 9.↵
Баррангу, Р., Э. Альтерманн, Р. Хаткинс, Р. Кано и Т. Р. Клаенхаммер. 2003. Функциональный и сравнительный анализ оперона, участвующего в утилизации фруктоолигосахаридов с помощью Lactobacillus acidophilus . Proc. Natl. Акад. Sci. USA100 : 8957-8962.
- 10.↵
Баррангу, Р., М. А. Азкарат-Перил, Т. Дуонг, С. Б. Коннерс, Р. М. Келли и Т. Р. Клаенхаммер. 2006. Глобальный анализ использования углеводов Lactobacillus acidophilus с использованием микрочипов кДНК.Proc. Natl. Акад. Sci. USA103 : 3816-3821.
- 11.↵
Begley, M., C.G. Gahan и C. Hill. 2005. Взаимодействие бактерий и желчи. FEMS Microbiol. Ред.29 : 625-651.
- 12.
Bellon-Fontaine, M.-N., J. Rault, and C. J. van Oss. 1996. Адгезия микробов к растворителям: новый метод определения электронодонорных / электроноакцепторных или кислотно-основных свойств Льюиса микробных клеток.Colloids Surf. B Биоинтерфейсы7 : 47-53.
- 13.↵
Бендцен, Дж. Д., Х. Нильсен, Г. фон Хейне и С. Брунак. 2004. Улучшенное предсказание сигнальных пептидов: SignalP 3.0. J. Mol. Biol.340 : 783-795.
- 14.↵
Битан-Банин Г., Р. Ортенберг и М. Мевареч. 2003. Разработка системы нокаута гена для галофильных архей Haloferax volcanii с использованием гена pyrE .J. Bacteriol.185 : 772-778.
- 15.↵
Боке, Дж. Д., Ф. Лакроут и Г. Р. Финк. 1984. Положительный отбор мутантов, лишенных активности оротидин-5′-фосфатдекарбоксилазы в дрожжах: устойчивость к 5-фтороротовой кислоте. Мол. Genet.197 : 345-346.
- 16.
Бут, Х. Дж., К. П. Колен, Б. Пот, К. Керстерс и П. Х. Поуэлс. 1996. Наличие двух генов, кодирующих белок S-слоя, сохраняется среди видов, родственных Lactobacillus acidophilus .Микробиология 142 : 2375-2384.
- 17.
Бут, Х. Дж., К. П. Колен и П. Х. Поуэлс. 1995. Идентификация, клонирование и нуклеотидная последовательность молчащего гена белка S-слоя Lactobacillus acidophilus ATCC 4356, который имеет большое сходство с геном белка S-слоя этого вида. J. Bacteriol.177 : 7222-7230.
- 18.↵
Бут, Х. Дж., К. П. Колен, Дж. М. ван Ноорт и П.H. Pouwels. 1993. Белок S-слоя Lactobacillus acidophilus ATCC 4356: очистка, экспрессия в Escherichia coli и нуклеотидная последовательность соответствующего гена. J. Bacteriol.175 : 6089-6096.
- 19.↵
Bruno-Barcena, J. M., M. A. Azcarate-Peril, T. R. Klaenhammer и H. M. Hassan. 2005. Безмаркерная хромосомная интеграция гена супероксиддисмутазы марганца ( sodA ) из Streptococcus thermophilus в Lactobacillus gasseri .FEMS Microbiol. Lett.246 : 91-101.
- 20.↵
Бак, Б. Л., Э. Альтерманн, Т. Свингеруд и Т. Р. Клаенхаммер. 2005. Функциональный анализ предполагаемых факторов адгезии у Lactobacillus acidophilus NCFM. Прил. Environ. Microbiol.71 : 8344-8351.
- 21.↵
Калланан, М., П. Калета, Дж. О’Каллаган, О. О’Салливан, К. Джордан, О. Маколифф, А. Санградор-Вегас, Л. Слэттери, GF Фицджеральд, Т.Бересфорд и Р. П. Росс. 2008. Последовательность генома Lactobacillus helveticus , организма, отличающегося селективной потерей гена и увеличением элемента последовательности вставки. J. Bacteriol. 190 : 727-735.
- 22.↵
Добсон, А. Э., Р. Б. Санозски-Дауэс и Т. Р. Клаенхаммер. 2007. Идентификация оперона и индуцирующего пептида, участвующих в продукции лактацина B Lactobacillus acidophilus . J. Appl. Microbiol.103 : 1766-1778.
- 23.↵
Duong, T., R. Barrangou, W. M. Russell и T. R. Klaenhammer. 2006. Характеристика локуса tre и анализ криопротекции трегалозы в Lactobacillus acidophilus NCFM. Прил. Environ. Microbiol.72 : 1218-1225.
- 24.↵
Fabret, C., S. D. Ehrlich, P. Noirot. 2002. Новая система доставки мутаций для подходов в масштабе генома в Bacillus subtilis .Мол. Microbiol.46 : 25-36.
- 25.↵
Фукагава, Т., Н. Хейворд, Дж. Ян, К. Аззалин, Д. Гриффин, А. Ф. Стюарт и У. Браун. 1999. Ген HPRT курицы: контрселективный маркер для линии клеток DT40. Nucleic Acids Res.27 : 1966-1969.
- 26.↵
Goh, Y.J., C. Zhang, A.K.Benson, V. Schlegel, J.H. Lee и R.W. Hutkins. 2006. Идентификация предполагаемого оперона, участвующего в утилизации фруктоолигосахаридов с помощью Lactobacillus paracasei .Прил. Environ. Microbiol.72 : 7518-7530.
- 27.↵
Hanahan, D. 1985. Методика трансформации E. coli , с. 109-135. В Д. М. Гловер (ред.), Клонирование ДНК: практический подход, т. 1. IRL Press Ltd., Оксфорд, Англия.
- 28.↵
Хортон, Р. М., Х. Д. Хант, С. Н. Хо, Дж. К. Пуллен и Л. Р. Пиз. 1989. Разработка гибридных генов без использования рестрикционных ферментов: сплайсинг генов путем перекрывающегося удлинения.Gene77 : 61-68.
- 29.↵
Якава-Вильянен, М., С. Авалл-Яаскелайнен, П. Месснер, У. Б. Слейтр и А. Палва. 2002. Выделение трех новых генов белка поверхностного слоя ( slp ) из Lactobacillus brevis ATCC 14869 и характеристика изменения их экспрессии в аэрированных и анаэробных условиях. J. Bacteriol. 184 : 6786-6795.
- 30.↵
Kimmel, S.A., и Р. Ф. Робертс. 1998. Разработка питательной среды, подходящей для продукции экзополисахаридов с помощью Lactobacillus delbrueckii spp. bulgaricus RR. Int. J. Food Microbiol. 40 : 87-92.
- 31.↵
Клаенхаммер, Т. Р., Р. Баррангу, Б. Л. Бак, М. А. Азкарат-Перил и Э. Альтерманн. 2005. Геномные особенности молочнокислых бактерий, влияющих на биотехнологию и здоровье. FEMS Microbiol. Ред.29 : 393-409.
- 32.↵
Константинов, С.Р., Х. Шмидт, В.М. де Вос, С.К. Бруйнс, С.К. Сингх, Ф. Валенс, Д. Молле, С. Лортал, Э. Альтерманн, Т. Р. Клаенхаммер и Ю. ван Койк. 2008. Белок S-слоя A Lactobacillus acidophilus NCFM регулирует функции незрелых дендритных клеток и Т-клеток. Proc. Natl. Акад. Sci. USA105 : 19474-19479.
- 33.↵
Кристич, К. Дж., Д. А. Маниас и Г. А. Данни. 2005. Разработка метода безмаркерного генетического обмена в Enterococcus faecalis и его использование при конструировании мутанта srtA . Прил. Environ. Microbiol.71 : 5837-5849.
- 34.↵
Law, J., G. Buist, A. Haandrikman, J. Kok, G. Venema, and K. Leenhouts. 1995. Система для генерации хромосомных мутаций в Lactococcus lactis , которая позволяет быстро анализировать целевые гены. J. Bacteriol.177 : 7011-7018.
- 35.↵
Maguin, E., P. Duwat, T. Hege, D. Ehrlich и A. Gruss. 1992. Новая термочувствительная плазмида для грамположительных бактерий. J. Bacteriol. 174 : 5633-5638.
- 36.↵
Мартинуссен, Дж., П. Глейзер, П. С. Андерсен и Х. Х. Саксилд. 1995. Два гена, кодирующие урацилфосфорибозилтрансферазу, присутствуют в Bacillus subtilis . J. Bacteriol. 177 : 271-274.
- 37.↵
Martinussen, J., and K. Hammer. 1994. Клонирование и характеристика upp , гена, кодирующего урацилфосфорибозилтрансферазу из Lactococcus lactis . J. Bacteriol.176 : 6457-6463.
- 38.↵
McAuliffe, O., R.J. Cano и T.R. Klaenhammer. 2005. Генетический анализ активности двух гидролаз желчных солей в Lactobacillus acidophilus NCFM. Прил. Environ. Microbiol.71 : 4925-4929.
- 39.↵
Мерритт, Дж., П. Цанг, Л. Чжэн, В. Ши и Ф. Ци. 2007. Создание основанной на контрселекции системы делеции в рамке считывания для генетических исследований Streptococcus mutans . Oral Microbiol. Immunol.22 : 95-102.
- 40.↵
Neuhard, J. 1982. Использование предварительно образованных пиримидиновых оснований и нуклеозидов, с. 95-148. В А. Мунк-Петерсен (ред.), Метаболизм нуклеотидов, нуклеозидов и азотистых оснований в микроорганизмах.Academic Press, Лондон, Соединенное Королевство.
- 41.↵
Peck, R. F., S. DasSarma, and M. P. Krebs. 2000. Нокаут гомологичного гена в архее Halobacterium salinarium с ura3 в качестве контрселективного маркера. Мол. Microbiol.35 : 667-676.
- 42.↵
Pfeiler, E. A., M. A. Azcarate-Peril и T. R. Klaenhammer. 2007. Характеристика нового индуцируемого желчью оперона, кодирующего двухкомпонентную регуляторную систему в Lactobacillus acidophilus .J. Bacteriol. 189 : 4624-4634.
- 43.
Pritchett, M. A., J. K. Zhang, W. W. Metcalf. 2004. Разработка метода безмаркерного генетического обмена для Methanosarcina acetivorans C2A и его использование в создании новых генетических инструментов для метаногенных архей. Прил. Environ. Microbiol.70 : 1425-1433.
- 44.↵
Рид, Дж. Л. и А. Коллмер. 1987. Картридж nptl-sacB-sacR для конструирования направленных немаркированных мутаций в грамотрицательных бактериях с помощью мутагенеза с заменой маркеров и вытеснением.Gene57 : 239-246.
- 45.↵
Rouillard, J. M., M. Zuker, and E. Gulari. 2003. OligoArray 2.0: дизайн олигонуклеотидных зондов для ДНК-микрочипов с использованием термодинамического подхода. Nucleic Acids Res.31 : 3057-3062.
- 46.↵
Russell, W. M., and T. R. Klaenhammer. 2001. Эффективная система для направленной интеграции в хромосомы Lactobacillus acidophilus и Lactobacillus gasseri посредством гомологичной рекомбинации.Прил. Environ. Microbiol.67 : 4361-4364.
- 47.↵
Sanders, M. E., and T. R. Klaenhammer. 2001. Научное обоснование функциональности Lactobacillus acidophilus NCFM как пробиотика. J. Dairy Sci. 84 : 319-331.
- 48.↵
Сара М. и У. Б. Слейтр. 2000. Белки S-слоя. J. Bacteriol. 182 : 859-868.
- 49.↵
Уэки, Т., С. Иноуэ и М. Иноуе. 1996. Положительно-отрицательные кассеты KG для конструирования мультигенных делеций с использованием одного лекарственного маркера. Gene183 : 153-157.
- 50.↵
van der Mei, H. C., B. van de Belt-Gritter, P. H. Pouwels, B. Martinez и H. J. Busscher. 2003. Гидрофобность клеточной поверхности передается белками S-слоя — исследование на рекомбинантных лактобациллах. Colloids Surf. B Биоинтерфейсы 28 : 127-134.
- 51.↵
Уокер, Д. К., К. Аояма и Т. Р. Клаенхаммер. 1996. Электротрансформация Lactobacillus acidophilus группы A1. FEMS Microbiol. Lett.138 : 233-237.
- 52.↵
Уокер, Д. К., и Т. Р. Кленхаммер. 1994. Выделение нового элемента вставки группы IS3 и конструирование вектора интеграции для Lactobacillus spp. J. Bacteriol.176 : 5330-5340.
- 53.№
Уолтер, Дж., П. Шанно, Г. В. Тэннок, Д. М. Лоуч, Ф. Даль Белло, Х. Ф. Дженкинсон, У. П. Хэммс и К. Хертель. 2005. Поверхностный белок с высокой молекулярной массой (Lsp) и метионинсульфоксидредуктаза B (MsrB) вносят свой вклад в экологические характеристики Lactobacillus reuteri в кишечнике мышей. Прил. Environ. Microbiol.71 : 979-986.
- 54.↵
Ван, Г., С. П. Кеннеди, С. Фасилудин, К. Ренсинг и С. ДасСарма. 2004. Устойчивость к мышьяку у Halobacterium sp. штамм NRC-1 исследовали с использованием улучшенной системы нокаута гена. J. Bacteriol.186 : 3187-3194.
- 55.↵
Wei, M.-Q., C.M. Rush, J.M. Norman, L.M. Hafner, R.J. Epping и P. Timms. 1995. Усовершенствованный метод трансформации штаммов Lactobacillus с использованием электропорации. J. Microbiol. Методы 21 : 97-109.
Mentor UPP
Центр карьеры
Не забудьте заглянуть на веб-страницу Career Services, чтобы узнать о предстоящих тренингах по профессиональному развитию и семинарах по стажировке.Это прекрасная возможность начать поиски карьеры.
Подробнее »Мэдлин Гилмер
Мэдлин Гилмер
Я знал с первого семинара, что Mentor UPP — это сообщество, которым я хотел бы окружить себя по мере прохождения студенческих лет. Я установил невероятные отношения и связи с преподавателями, выпускниками, профессорами и другими студентами, приняв участие в этой организации. Существует множество наставников, которые вольются в вас как в студента и члена сообщества.Я так благодарен за Mentor UPP, и я всегда говорю об этом другим студентам-инженерам из Университета Алабамы.
Мэдлин Гилмер, младший
Алора Терри
Алора Терри
Наставник UPP был для меня поистине бесценным опытом. Это дало мне возможность узнать больше о нетворкинге и познакомиться с другими людьми из инженерного колледжа, одновременно получая лидерский опыт в качестве наставника. Членство в Консультативном совете позволило мне расти как молодому профессионалу и укрепило мою уверенность в себе.Кроме того, у меня была возможность завести друзей на всю жизнь, с которыми я надеюсь поддерживать связь после окончания учебы.
Алора Терри, выпускница
Саманта Томас
Сэм Томас
Наставник UPP был потрясающим опытом. Я стал наставником в младших классах и очень сблизился со своими подопечными. Я посетил большинство мероприятий и очень быстро завел новых друзей и знакомых в инженерном колледже. Вскоре после этого я присоединился к консультативному совету и смог озвучить перемены и поддержать эту программу, которая мне понравилась.Мне пришлось работать бок о бок с Нэнси Холмс и Гейл Хауэлл, двумя потрясающими и вдохновляющими женщинами, которые ценили мой вклад и поддерживали меня во всех моих академических начинаниях. Я смог вырасти и развить свои лидерские качества в обстановке, где я чувствовал себя ценным и услышанным. Я рекомендую эту программу каждому инженеру, которого встречаю, и мне так, так, так грустно, что я ухожу. Я надеюсь на лучшее для программы и буду петь ей дифирамбы, куда бы я ни пошел.
Саманта Томас, выпускница
Клинт Смит
Клинт Смит
Я был частью Mentor UPP в течение пяти лет.С первого года обучения я знал, что эта организация позволит мне расти как личности, в академическом, профессиональном и умственном плане. Программа Mentor UPP позволила мне встретиться с людьми из всего инженерного колледжа, и отношения, которые я установил в рамках программы, будут длиться вечно. Передо мной стояла задача расти еще больше и быть лучшим учеником и человеком. Эта программа изменила мой опыт учебы в колледже к лучшему. Мой единственный совет будущим студентам: чем больше вы отдадите себя организации (не только этой, но и любой другой), тем больше преимуществ и опыта вы получите.
Клинт Смит, выпускник
Блэр Батлер
Блэр Батлер
Наставник UPP был для меня таким потрясающим опытом. Он научил меня лидерским качествам, навыкам работы в сети и помог мне установить контакты с другими студентами и выпускниками инженерного колледжа. Фактически, связи, которые я установил через Mentor UPP, привели к тому, что я получил работу после окончания учебы! Я не могу порекомендовать эту программу студентам любого уровня.
Блэр Батлер, выпускница
Кристин Харрис
Кристин Харрис
Благодаря этой программе я не только смог познакомиться со студентами из разных дисциплин, но и подружиться.Эта программа дала мне возможность быть наставником и общаться с другими студентами-инженерами. Членство в консультативном совете также дало мне возможность работать с другими членами, чтобы помочь сформировать направление, в котором движется программа. Это прекрасная возможность, и я рад, что поступил на первый год обучения.
Кристин Харрис, старший
.