Ассемблер для Raspberry Pi. Практическое руководство (Брюс Смит)
1 280 ₽
1 079 ₽
+ до 192 баллов
Бонусная программа
Итоговая сумма бонусов может отличаться от указанной, если к заказу будут применены скидки.
Купить
Цена на сайте может отличаться от цены в магазинах сети. Внешний вид книги может отличаться от изображения на сайте.
В наличии 2 шт
В наличии в 128 магазинах. Смотреть на карте
2
Цена на сайте может отличаться от цены в магазинах сети. Внешний вид книги может отличаться от изображения на сайте.
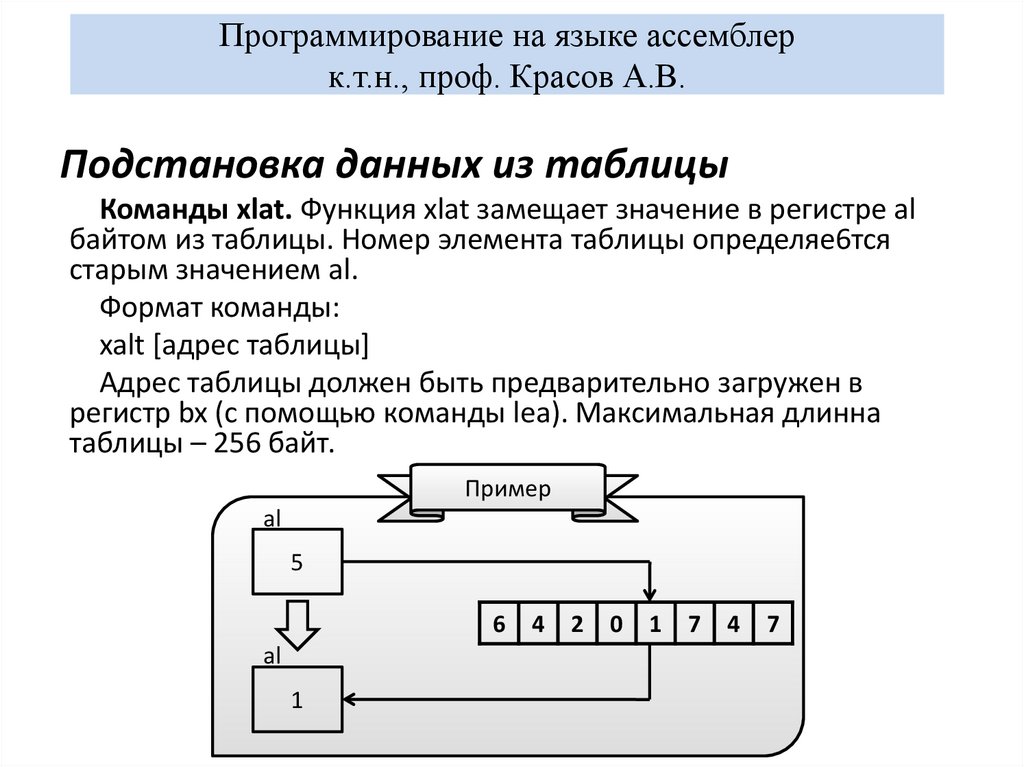
Рассмотрены основы программирования на языке ассемблера для процессоров ARM на примере Raspberry Pi с операционной системой Raspberry Pi OS. Приведены подробные сведения об архитектуре и особенностях ARM, вызовах операционной системы.
Электронный архив на сайте издательства содержит исходный код программ из книги.
Описание
Характеристики
Рассмотрены основы программирования на языке ассемблера для процессоров ARM на примере Raspberry Pi с операционной системой Raspberry Pi OS. Приведены подробные сведения об архитектуре и особенностях ARM, вызовах операционной системы. Подробно описан синтаксис ассемблера для ARM. Рассмотрены компоновщик GCC, отладка с GDB, использование функций языка C в ассемблере с помощью библиотеки libc. Описаны функции GPIO, система команд ARM Neon и команды Thumb. Все разделы снабжены практическими примерами.
Электронный архив на сайте издательства содержит исходный код программ из книги.
БХВ
На товар пока нет отзывов
Поделитесь своим мнением раньше всех
Как получить бонусы за отзыв о товаре
1
Сделайте заказ в интернет-магазине
2
Напишите развёрнутый отзыв от 300 символов только на то, что вы купили
3
Дождитесь, пока отзыв опубликуют.
Если он окажется среди первых десяти, вы получите 30 бонусов на Карту Любимого Покупателя. Можно писать
неограниченное количество отзывов к разным покупкам – мы начислим бонусы за каждый, опубликованный в
первой десятке.
Правила начисления бонусов
Если он окажется среди первых десяти, вы получите 30 бонусов на Карту Любимого Покупателя. Можно писать неограниченное количество отзывов к разным покупкам – мы начислим бонусы за каждый, опубликованный в первой десятке.
Правила начисления бонусов
Книга «Ассемблер для Raspberry Pi. Практическое руководство» есть в наличии в интернет-магазине «Читай-город» по привлекательной цене.
Если вы находитесь в Москве, Санкт-Петербурге, Нижнем Новгороде, Казани, Екатеринбурге, Ростове-на-Дону или любом
другом регионе России, вы можете оформить заказ на книгу
Брюс Смит
«Ассемблер для Raspberry Pi. Практическое руководство» и выбрать удобный способ его получения: самовывоз, доставка курьером или отправка
почтой. Чтобы покупать книги вам было ещё приятнее, мы регулярно проводим акции и конкурсы.
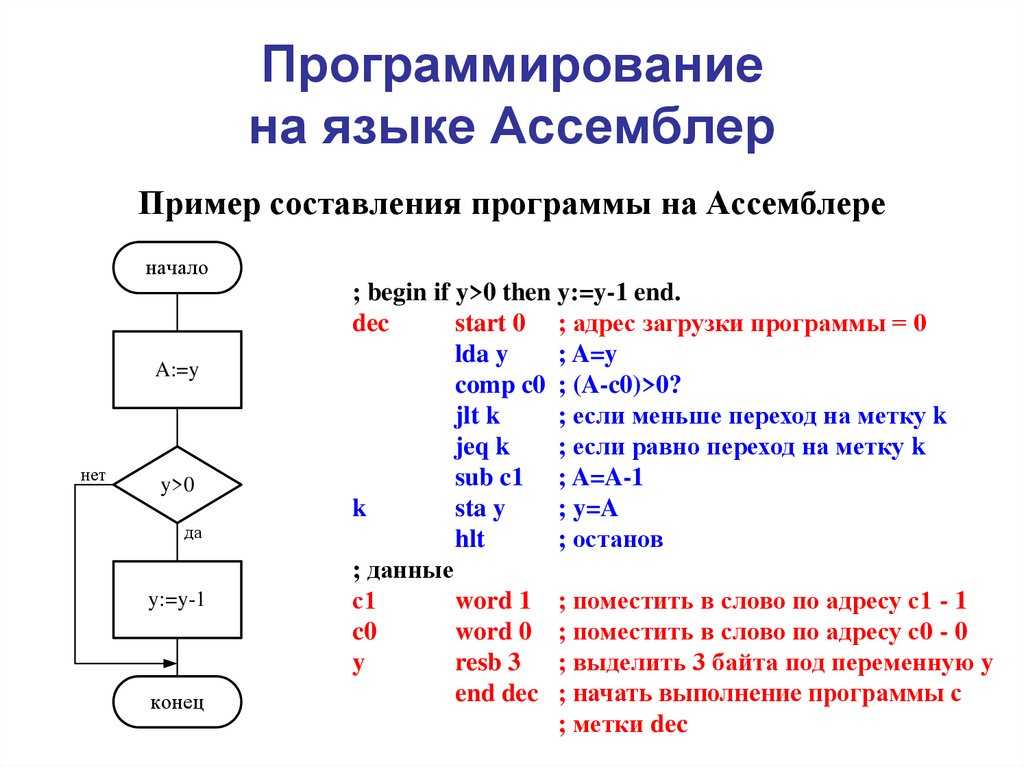
Научно-образовательный портал ТУСУР | Основы программирования на языке Ассемблера: Лабораторный практикум / Кирнос В. Н. — 2007. 106 с.
Лабораторный практикум
ВВЕДЕНИЕ
ОПЕРАТИВНАЯ ПАМЯТЬ
РЕГИСТРЫ
Регистры общего назначения
Сегментные регистры
Указатель команд
ПРЕДСТАВЛЕНИЕ КОМАНД
§1. ВЫЧИСЛЕНИЕ АРИФМЕТИЧЕСКИХ ВЫРАЖЕНИЙ
1.1 ПОНЯТИЕ ОБ АРИФМЕТИЧЕСКИХ ОПЕРАЦИЯХ
1.2 ОФОРМЛЕНИЕ ПРОГРАММЫ НА АССЕМБЛЕРЕ
1.3 ИСПОЛНЕНИЕ ПРОГРАММЫ
Практические задания к § 1
§ 2. ПЕРЕХОДЫ И ВЕТВЛЕНИЯ НА АССЕМБЛЕРЕ
Практические задания к §2
§ 3 ЦИКЛЫ СО СЧЕТЧИКОМ
Практические задания к § 3
§4. МАССИВЫ
4.1 ОДНОМЕРНЫЕ МАССИВЫ
4.2 ДВУМЕРНЫЕ МАССИВЫ
Практические задания к §4
§5. ПРОЦЕДУРЫ
5.1 ПОНЯТИЕ О ПРОЦЕДУРЕ
5.2 ПЕРЕДАЧА ПАРАМЕТРОВ В ПРОЦЕДУРУ
5.3 РЕКУРСИВНЫЕ ПРОЦЕДУРЫ
Практические задания к §5
§ 6.
Кафедра комплексной информационной безопасности электронно-вычислительных систем
Автор: Кирнос В. Н.
Год издания: 2007
Количество страниц: 106
Скачиваний: 17
Оглавление (содержание)
В пособии кратко рассматривается основы программирования на Ассемблере для процессора 8086. Во введении даются краткие теоретические сведения по организации компьютера. Далее последовательно рассматривается реализация основных типов алгоритмических структур (линейная, разветвленная, циклическая), работа с массивами и функциями на данном языке. Затем особое внимание уделяется организации ввода и вывода на языке ассемблера.
Во введении даются краткие теоретические сведения по организации компьютера. Далее последовательно рассматривается реализация основных типов алгоритмических структур (линейная, разветвленная, циклическая), работа с массивами и функциями на данном языке. Затем особое внимание уделяется организации ввода и вывода на языке ассемблера.
Дополнительно рассматривается работа с файлами и с вещественными числами на Ассемблере.
В конце каждого параграфа даются практические задания для самостоятельного выполнения (их следует использовать при выполнении лабораторных работ).
Пособие используется для изучения 1 дисциплины
Информационные системы в экономике
10.05.04 Информационно-аналитические системы безопасности (Информационная безопасность финансовых и экономических структур) Очная форма обучения, план набора 2012 г. План в архиве
Похожие пособия
Архитектура вычислительных систем: Методические указания по выполнению лабораторных работ и организации самостоятельной работы / Гриценко Ю.
 Б. — 2017. 30 с.
Б. — 2017. 30 с. Системное программное обеспечение: Учебное пособие / Гриценко Ю. Б. — 2006. 174 с.
Микропроцессорные устройства и системы: Методические указания по проведению лабораторных работ / Антипин М. Е. — 2012. 13 с.
Архитектура вычислительных систем: Методические указания по выполнению лабораторных работ и организации самостоятельной работы для студентов направления подготовки Программная инженерия (квалификация (степень) «бакалавр») / Гриценко Ю. Б. — 2016. 73 с.
Архитектура вычислительных систем, аппаратное и программное обеспечение: Методические указания по выполнению лабораторных работ и организации самостоятельной работы для студентов направления подготовки Бизнес-информатика (квалификация (степень) «бакалавр») / Гриценко Ю. Б. — 2016. 74 с.
Учебное пособие по сборкеDe Novo | Geneious Prime
Общие рекомендации по сборке de novo Сборка De novo — одна из самых ресурсоемких задач с оперативной и процессорной памятью, которые можно выполнять в Geneious Prime. Обзор требований к оборудованию для сборки de novo с использованием ассемблера Geneious de novo см. в нашей публикации базы знаний.
Обзор требований к оборудованию для сборки de novo с использованием ассемблера Geneious de novo см. в нашей публикации базы знаний.
Для улучшения качества сборки и сокращения времени и оперативной памяти, необходимых для сборки, всегда необходимо:
1. Обрезать концы с помощью BBDuk. Строго обрежьте со значением «Q» 20 или выше.
2. Стремитесь к монтажному покрытию в 50-100 раз. Более высокое покрытие обычно не улучшает конечные результаты и увеличивает требования к оперативной памяти и время сборки. См. следующую ссылку для получения информации о том, как рассчитать ожидаемое покрытие на основе ожидаемого размера генома, средней длины чтения (обрезанного) и количества прочтений.
Чтобы de novo собрать подмножество ваших ридов, используйте один из следующих вариантов:
а) В окне настроек сборки De novo установите флажок Использовать X% данных и установите соответствующее значение на основе расчетов покрытия. Это всегда будет использовать первые X% чтений в вашем списке.
Это всегда будет использовать первые X% чтений в вашем списке.
б) Используйте Рабочие процессы → Случайные выборки последовательностей , чтобы создать случайно выбранное подмножество (попарно) вашего списка для чтения.
c) Используйте нормализацию, чтобы уменьшить размер набора данных, как описано ниже.
Воля de novo 9Сборка 0005 дает мне полный непрерывный геном?
Все алгоритмы сборки de novo , независимо от используемой методики, не могут однозначно выполнять сборку по идеальным повторам, если длина повторяющейся единицы превышает длину считывания или размер вставки парного считывания. На практике это означает, что сборка геномных данных короткого считывания Illumina в большинстве случаев приведет к созданию множества непересекающихся контигов.
Например, микробные геномы обычно содержат несколько копий кластера генов рРНК, каждая из которых содержит почти идеальные повторы генов SSU (16S), LSU (23S) и 5S рРНК. Эти идентичные кластеры рРНК будут препятствовать сборке и восстановлению единой непрерывной консенсусной последовательности. В большинстве случаев другие повторяющиеся единицы (дублированные гены, транспозоны и т. д.) также будут генерировать дальнейшие «разрывы» в de novo в сборе.
Эти идентичные кластеры рРНК будут препятствовать сборке и восстановлению единой непрерывной консенсусной последовательности. В большинстве случаев другие повторяющиеся единицы (дублированные гены, транспозоны и т. д.) также будут генерировать дальнейшие «разрывы» в de novo в сборе.
Все чтения, полученные из повторов, в конечном итоге будут собраны вместе в конце одного контига. Как следствие, покрытие области собранных повторов будет выше, чем среднее покрытие уникальной части генома.
Другие инструменты предварительной обработкиИсправление ошибок и нормализация показаний
Доступ через меню Последовательность → Исправление ошибок и нормализация показаний.
Инструмент «Исправить ошибки и нормализовать считывания» использует BBNorm, который предназначен для нормализации покрытия сборки за счет уменьшения выборки чтений в областях с большой глубиной генома, что приводит к более равномерному распределению покрытия. Важно отметить, что нормализация не удалит чтения в областях с более низким покрытием.
Важно отметить, что нормализация не удалит чтения в областях с более низким покрытием.
Нормализация может существенно уменьшить размер набора данных, а впоследствии для сборки de novo сократить время сборки и снизить требования к ОЗУ.
Обратите внимание, что нормализация потенциально может «усилить» ошибки в «сложных для последовательности» областях до такой степени, что ошибки будут казаться значительными. Поэтому, если вы используете нормализацию, мы рекомендуем использовать стратегию, изображенную на блок-схеме, показанной ниже. Эта комбинированная стратегия нормализации/сборки de novo /сопоставления со справочной стратегией обычно будет намного быстрее, чем попытка de novo сборка вашего полного набора данных.
Объединить парные чтения
Доступ через меню Последовательность → Объединить парные чтения
Этот инструмент использует BBMerge и предназначен для объединения двух перекрывающихся парных чтений в одно чтение. Этот инструмент полезен для получения консенсуса на основе перекрывающихся чтений, полученных при секвенировании ампликонов.
Этот инструмент полезен для получения консенсуса на основе перекрывающихся чтений, полученных при секвенировании ампликонов.
Удалить повторяющиеся чтения
Доступ через меню Последовательность → Удалить повторяющиеся чтения
Этот инструмент использует Dedupe и предназначен для поиска и удаления всех содержащихся и перекрывающихся последовательностей в наборе данных.
Удалить химер
Доступ через меню Последовательность → Удалить химерные чтения.
Этот инструмент будет фильтровать химерные чтения из данных секвенирования путем сравнения с эталонной базой данных. Вы можете выбрать между объединенным общедоступным алгоритмом UCHIME или загрузить и использовать более быстрый USEARCH 8. Обратите внимание, что бесплатная версия USEARCH 8 ограничена использованием 4 ГБ ОЗУ и поэтому не может обрабатывать большие наборы данных NGS.
Разделение штрих-кода
Доступ через меню Последовательность → Разделение по штрих-кодам.
Этот инструмент демультиплексирует пользовательские данные со штрих-кодом в отдельные списки. Инструмент имеет 454 предустановки штрих-кода MID, или вы можете определить и использовать свои собственные наборы штрих-кодов.
Примечание: Демультиплексирование всегда следует выполнять перед обрезкой с помощью BBduk.
Практические подходы к фолдингу и сборке белков: спектроскопические стратегии в термодинамике и кинетике
Сохранить цитату в файл
Формат: Резюме (текст) PubMedPMIDAbstract (текст) CSV
Добавить в коллекции
- Создать новую коллекцию
- Добавить в существующую коллекцию
Назовите свою коллекцию:
Имя должно содержать менее 100 символов
Выберите коллекцию:
Не удалось загрузить вашу коллекцию из-за ошибки
Повторите попытку
Добавить в мою библиографию
- Моя библиография
Не удалось загрузить делегатов из-за ошибки
Повторите попытку
Ваш сохраненный поиск
Название сохраненного поиска:
Условия поиска:
Тестовые условия поиска
Электронная почта: (изменить)
Который день? Первое воскресеньеПервый понедельникПервый вторникПервая средаПервый четвергПервая пятницаПервая субботаПервый деньПервый рабочий день
Который день? ВоскресеньеПонедельникВторникСредаЧетвергПятницаСуббота
Формат отчета: РезюмеРезюме (текст)АбстрактАбстракт (текст)PubMed
Отправить максимум:
1 шт.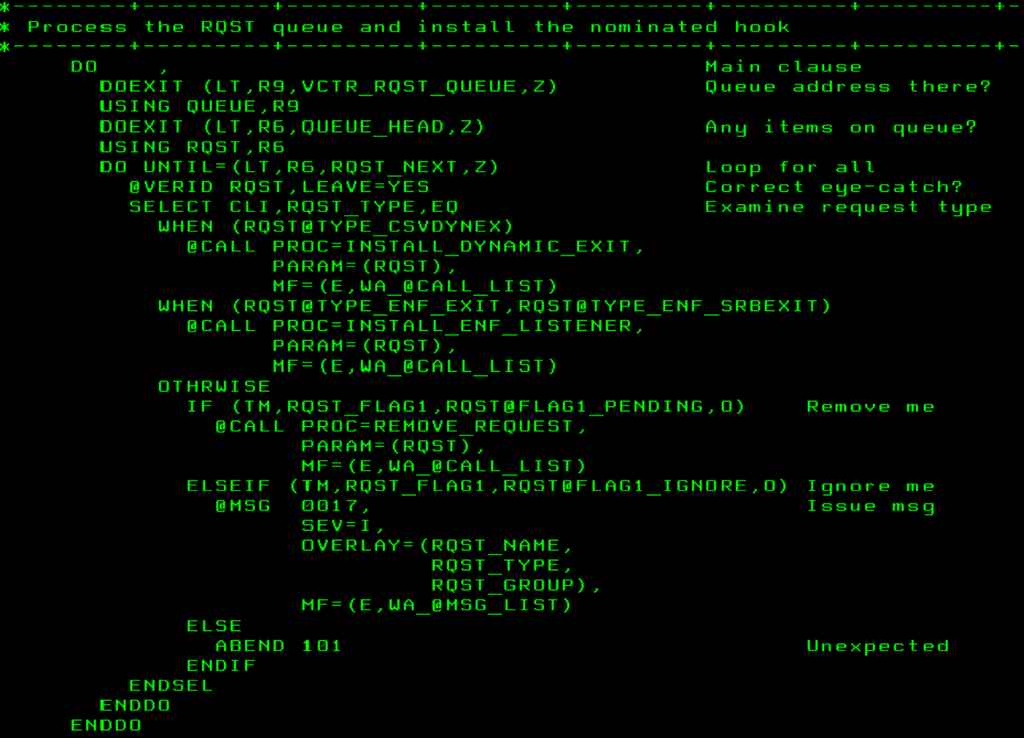 5 шт. 10 шт. 20 шт. 50 шт. 100 шт. 200 шт.
5 шт. 10 шт. 20 шт. 50 шт. 100 шт. 200 шт.
Отправить, даже если нет новых результатов
Необязательный текст в электронном письме:
Создайте файл для внешнего программного обеспечения для управления цитированием
doi: 10.1016/S0076-6879(08)04201-8.
Джад Уолтерс 1 , Сара Л. Милам, A Clay Clark
принадлежность
- 1 Кафедра молекулярной и структурной биохимии, Государственный университет Северной Каролины, Роли, Северная Каролина, США.
- PMID: 19289201
- PMCID: PMC2778058
- DOI:
10.
 1016/С0076-6879(08)04201-8
1016/С0076-6879(08)04201-8
Бесплатная статья ЧВК
Джад Уолтерс и др. Методы Энзимол. 2009.
Бесплатная статья ЧВК
. 2009; 455:1-39.
doi: 10.1016/S0076-6879(08)04201-8.
Авторы
Джад Уолтерс 1 , Сара Л. Милам, А. Клэй Кларк
принадлежность
- 1 Кафедра молекулярной и структурной биохимии, Государственный университет Северной Каролины, Роли, Северная Каролина, США.
- PMID: 19289201
- PMCID: PMC2778058
- DOI:
10.
 1016/С0076-6879(08)04201-8
1016/С0076-6879(08)04201-8
Абстрактный
Мы описываем здесь использование нескольких спектроскопий, таких как испускание флуоресценции, круговой дихроизм и дифференциальное тушение акриламидом, при изучении равновесия и кинетической укладки белков. Первый раздел, посвященный методам установления равновесия, предоставляет практическую информацию для определения конформационной стабильности белка. Кроме того, обсуждаются несколько моделей равновесной укладки, от мономера с двумя состояниями до гомодимера с четырьмя состояниями, что обеспечивает всеобъемлющий протокол для интерпретации кривых сворачивания. Во втором разделе основное внимание уделяется планированию эксперимента и интерпретации кинетических данных, таких как анализ фазы взрыва и экспоненциальные подборы, используемые для выяснения путей кинетического фолдинга. Кроме того, программы моделирования обычно используются для поддержки моделей складывания, созданных в результате кинетических экспериментов, и рассматриваются основы моделирования.
Цифры
Рисунок 1.1
(A) Спектры излучения после возбуждения…
Рисунок 1.1
(A) Спектры эмиссии после возбуждения при 280 нм. Данные для 0 M (○)…
Рисунок 1.1 (A) Спектры эмиссии после возбуждения при 280 нм. Данные для 0 9Показаны мочевина 0261 M (○) и 8 M (□). В этом примере пунктирная линия указывает длину волны, при которой можно отслеживать разворачивание из-за большой разницы в сигнале между нативным и развернутым белком. (B) Кривая развертывания равновесия, контролируемая флуоресцентным излучением при 320 нм (возбуждение при 280 нм). Указаны предпереходная, переходная и постпереходная области. (C) Нормализованные данные, демонстрирующие три зонда, использованные в экспериментах по разворачиванию. Развертывание контролировали по испусканию флуоресценции после возбуждения при 280 нм (○) или 295 нм (□) или по КД (Δ). Рефолдинговый белок (●) демонстрирует обратимость. (D) Несовпадение кривых развертывания при наблюдении с помощью различных спектроскопических методов, что предполагает более сложный механизм складывания, чем модель с двумя состояниями, предложенная одним методом. (E) Пример кривой равновесия с тремя состояниями. Сплошные линии на панелях C-E представляют соответствие данных модели развертывания мономера с двумя состояниями (панели C и D) или с тремя состояниями (панель E), как описано в тексте.
Развертывание контролировали по испусканию флуоресценции после возбуждения при 280 нм (○) или 295 нм (□) или по КД (Δ). Рефолдинговый белок (●) демонстрирует обратимость. (D) Несовпадение кривых развертывания при наблюдении с помощью различных спектроскопических методов, что предполагает более сложный механизм складывания, чем модель с двумя состояниями, предложенная одним методом. (E) Пример кривой равновесия с тремя состояниями. Сплошные линии на панелях C-E представляют соответствие данных модели развертывания мономера с двумя состояниями (панели C и D) или с тремя состояниями (панель E), как описано в тексте.
Рисунок 1.2
(A) Пример равновесия-развертывания…
Рисунок 1.2
(A) Пример кривой равновесия-развертывания для димерного белка, которая следует за…
Рисунок 1. 2
2 (A) Пример кривой равновесия-разворачивания для димерного белка, которая соответствует модели развертывания с двумя состояниями. Зависимость от концентрации белка демонстрируется увеличением средней точки перехода при увеличении концентрации белка (○ > □ > Δ). (B) Пример модели разворачивания димера с тремя состояниями, в которой зависимость концентрации белка наблюдается при первом переходе, демонстрируя присутствие мономерного промежуточного соединения. (C) Пример модели разворачивания димера с тремя состояниями, в которой зависимость концентрации белка наблюдается во втором переходе, демонстрируя присутствие димерного промежуточного соединения. (D) Пример модели разворачивания димера с четырьмя состояниями. (I) В этом случае середина первого перехода одинакова для каждой концентрации белка, а середина второго перехода увеличивается с увеличением концентрации белка (○ > ◇). Относительный сигнал, наблюдаемый между 3 M и 5 M мочевины также увеличиваются с увеличением концентрации белка, демонстрируя процесс развертывания с четырьмя состояниями, как описано в тексте.
Рисунок 1.3
(A) Гипотетический график флуоресценции…
Рисунок 1.3
(A) Гипотетический график сигнала флуоресценции в присутствии 0,7 M акриламид…
Рисунок 1.3 (A) Гипотетический график зависимости сигнала флуоресценции в присутствии 0,7 М акриламида от времени для реакции рефолдинга. Показаны сигналы развернутого (+) и нативного (x) белка в присутствии акриламида. Показаны данные для нескольких конечных концентраций мочевины (Δ, ◇, □, ○). (B) Гипотетический график сигнала эмиссии флуоресценции в зависимости от времени для реакции повторной укладки. Показаны сигналы развернутого (■) и нативного (●) белка. Конечная концентрация мочевины 3,45· M (○), 4,36 M (□), 5,27 M (◇) и 6,18 M (Δ).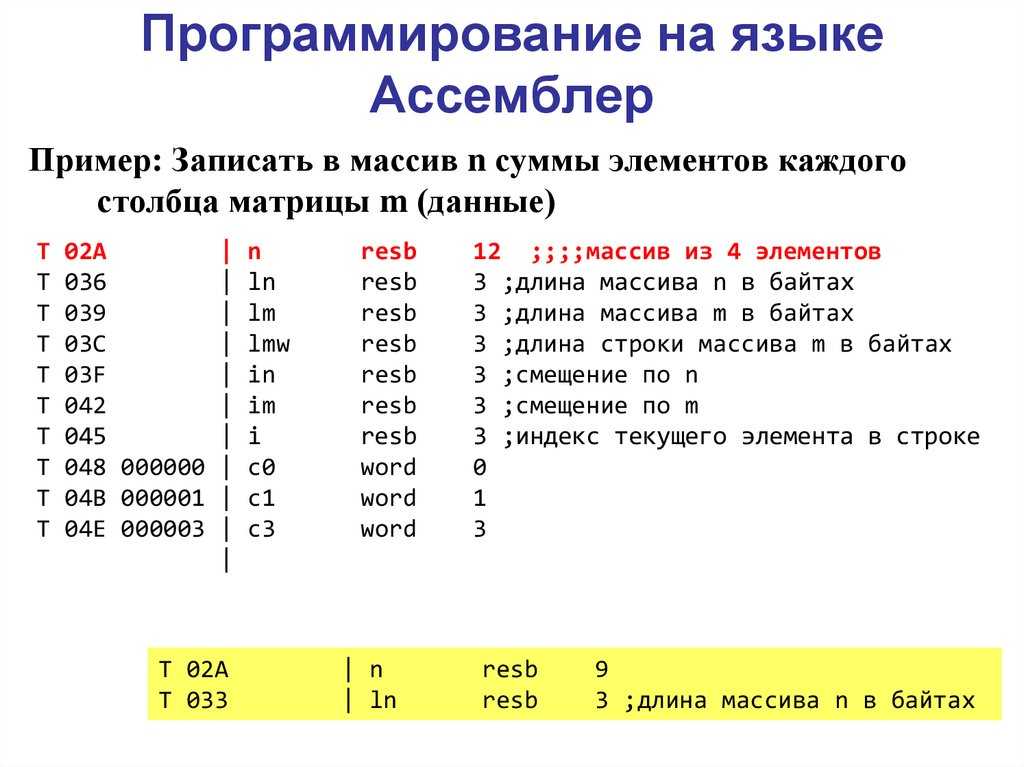 Область в рамке отмечает время от 1 до 5 мс. Стрелки показывают экстраполяцию сигнала к нулевому времени. (C) График сигнала фазы взрыва в зависимости от конечной концентрации мочевины. Непрерывная линия соответствует модели равновесного складывания с двумя состояниями, как описано в тексте.
Область в рамке отмечает время от 1 до 5 мс. Стрелки показывают экстраполяцию сигнала к нулевому времени. (C) График сигнала фазы взрыва в зависимости от конечной концентрации мочевины. Непрерывная линия соответствует модели равновесного складывания с двумя состояниями, как описано в тексте.
Рисунок 1.4
4.36 M данные перескладывания…
Рисунок 1.4
Данные повторной складывания 4,36 M из рис. 1.3B были приспособлены к одному…
Рисунок 1.4 Данные рефолдинга 4,36 M из рис. 1.3B соответствовали одному (A), двум (B) или трем (C) экспоненциальным уравнениям. Остатки посадок показаны под каждой панелью. (A) Одна экспоненциальная подгонка данных обеспечивает амплитуду 0,22 и скорость 33 с -1 . (B) Две экспоненциальные подгонки данных обеспечивают амплитуды 0,20 и 0,08 и скорость 50 и 0,3 сек -1 , соответственно. (C) Трехэкспоненциальная подгонка данных обеспечивает амплитуды 0,2, 0,08 и 0,05 и скорости 50, 0,3 и 3,3 × 10 -7 сек -1 соответственно.
(B) Две экспоненциальные подгонки данных обеспечивают амплитуды 0,20 и 0,08 и скорость 50 и 0,3 сек -1 , соответственно. (C) Трехэкспоненциальная подгонка данных обеспечивает амплитуды 0,2, 0,08 и 0,05 и скорости 50, 0,3 и 3,3 × 10 -7 сек -1 соответственно.
Рисунок 1.5
(A) Гипотетический шевронный участок…
Рисунок 1.5
(A) Гипотетический шевронный график кажущихся скоростей рефолдинга и разворачивания в зависимости от…
Рисунок 1.5 (A) Гипотетический шевронный график кажущейся скорости повторной укладки и разворачивания в зависимости от конечной концентрации мочевины. Сплошная линия соответствует модели кинетического складывания с двумя состояниями (Ferguson et al. , 1999) со следующими параметрами: kfh3O=200 с-1, kUh3O=0,015 с-1, м N-TS = 0,7, m U-TS = 0,6, Δ G H 2 O = 5,62 ккал/моль, m-значение = 1,3 ккал/моль/моль, мочевина{12}= 4,3М. (B) Пример последовательного пути с одним промежуточным продуктом, записанным в текстовом формате для KINSIM. X1, X2 и X3 — коэффициенты экстинкции U, I и N соответственно. (C) Гипотетический пример реакции рефолдинга 10 мкМ белка в течение 10 с. Популяции видов, показанные сплошными линиями, обозначены как U, I, N и U + I + N. Скорость перехода от U к I и от I к N составляет 1 и 0,3 с9.0183 -1 соответственно. Значения X1, X2 и X3 равны 0,01, 0,05 и 0,06 соответственно.
, 1999) со следующими параметрами: kfh3O=200 с-1, kUh3O=0,015 с-1, м N-TS = 0,7, m U-TS = 0,6, Δ G H 2 O = 5,62 ккал/моль, m-значение = 1,3 ккал/моль/моль, мочевина{12}= 4,3М. (B) Пример последовательного пути с одним промежуточным продуктом, записанным в текстовом формате для KINSIM. X1, X2 и X3 — коэффициенты экстинкции U, I и N соответственно. (C) Гипотетический пример реакции рефолдинга 10 мкМ белка в течение 10 с. Популяции видов, показанные сплошными линиями, обозначены как U, I, N и U + I + N. Скорость перехода от U к I и от I к N составляет 1 и 0,3 с9.0183 -1 соответственно. Значения X1, X2 и X3 равны 0,01, 0,05 и 0,06 соответственно.
Рисунок 1.6
Один последовательный (А) и два…
Рисунок 1. 6
6
Один последовательный (A) и два параллельных пути складывания (B и C), используемые в…
Рисунок 1.6 Один последовательный (A) и два параллельных пути складывания (B и C), используемые в моделировании KINSIM. (A) Развернутый вид складывается через две промежуточные формы, прежде чем сформировать нативный вид. (B) Два развернутых и местных вида существуют и взаимопревращаются. (C) Существуют три развернутых вида, где U 1 и U 2 сворачиваются в нативное состояние, а U 3 взаимопревращается с U 1 или U 2 . (D-F) Графики реакции рефолдинга Apaf-1 CARD в 3,64 М мочевины. Моделирование KINSIM для каждого участка показано сплошной линией. Для пути А концентрация I 1 было 10 мкМ . Коэффициент экстинкции для U, I 1 , I 2 и N составил 0, 0,0007, 0,0058 и 0,0063 соответственно. Скорости для I 1 до I 2 и I 2 до N составляли 45 и 0,6 с -1 соответственно. Для пути B концентрации U 1 и U 2 составляли 1 и 9 мкМ соответственно. Коэффициенты экстинкции для N 1 и N 2 составили 0,0074 и 0,0064. Ставки U 9От 0383 1 до N 1 , U 1 до U 2 , U 2 до N 2 , и N 1 до N 2 , и N 1 до N 2 , 70383 1 до N 2 , 50, 50, 50, до N 2 , 50, 50, и N 2 . -1 соответственно. Для пути C концентрации U 1 , U 2 и U 3 составляли 0,7, 8,3 и 1 мкМ . Коэффициент экстинкции N составил 0,0086. Курсы U 1 к N, U 2 к N и U 3 к U 1 или U 2 были 5000, 37 (скорость возврата 13) и 0,6 (скорость возврата 0,01) в секунду.
Для пути B концентрации U 1 и U 2 составляли 1 и 9 мкМ соответственно. Коэффициенты экстинкции для N 1 и N 2 составили 0,0074 и 0,0064. Ставки U 9От 0383 1 до N 1 , U 1 до U 2 , U 2 до N 2 , и N 1 до N 2 , и N 1 до N 2 , 70383 1 до N 2 , 50, 50, 50, до N 2 , 50, 50, и N 2 . -1 соответственно. Для пути C концентрации U 1 , U 2 и U 3 составляли 0,7, 8,3 и 1 мкМ . Коэффициент экстинкции N составил 0,0086. Курсы U 1 к N, U 2 к N и U 3 к U 1 или U 2 были 5000, 37 (скорость возврата 13) и 0,6 (скорость возврата 0,01) в секунду.
См. это изображение и информацию об авторских правах в PMC
Похожие статьи
Атомистическое описание сворачивания димерного белка.
Пиана С., Линдорф-Ларсен К., Шоу, Д.Е. Пиана С. и др. J Phys Chem B. 24 октября 2013 г.; 117 (42): 12935-42. дои: 10.1021/jp4020993. Epub 2013 15 августа. J Phys Chem B. 2013. PMID: 23882999
На пути к последовательному моделированию термодинамической и кинетической кооперативности белков: насколько применима картина переходного состояния к складыванию и разворачиванию?
Кая Х., Чан Х.С. Кая Х и др. Дж Мол Биол. 2002 25 января; 315 (4): 899-909. doi: 10.1006/jmbi.2001.5266. Дж Мол Биол. 2002. PMID: 11812156
Сворачивание цитохрома с лошади в восстановленном состоянии.
Бхуян А.К., Удгаонкар Дж.Б. Бхуян А.К. и соавт. Дж Мол Биол. 2001 г., 5 октября; 312(5):1135-60.
 doi: 10.1006/jmbi.2001.4993.
Дж Мол Биол. 2001.
PMID: 11580255
doi: 10.1006/jmbi.2001.4993.
Дж Мол Биол. 2001.
PMID: 11580255Использование нескольких спектроскопических методов для мониторинга равновесного разворачивания белков.
Эфтинк МР. Эфтинк МР. Методы Энзимол. 1995; 259:487-512. doi: 10.1016/0076-6879(95)59058-7. Методы Энзимол. 1995. PMID: 8538469Аннотация недоступна.
Спектроскопические исследования фолдинга белков: линейные и нелинейные методы.
Серрано А.Л., Вагеле М.М., Гай Ф. Серрано А.Л. и соавт. Белковая наука. 2012 февраля; 21 (2): 157-70. doi: 10.1002/pro.2006. Epub 2011 28 декабря. Белковая наука. 2012. PMID: 22109973 Бесплатная статья ЧВК. Обзор.
Посмотреть все похожие статьи
Цитируется
Сравнение складчатых ландшафтов эволюционно дивергентной прокаспазы-3.

Яо Л., Кларк А.С. Яо Л. и др. Представитель Biosci, 30 июня 2022 г.; 42(6):BSR20220119. дои: 10.1042/BSR20220119. Представитель Biosci, 2022 г. PMID: 35670809 Бесплатная статья ЧВК.
Характеристика циклофилина из Thaumarchaeota Nitrosopumilus maritimus: Влияние на разнообразие шапероноподобной активности в домене архей.
Кошик В., Анчал, Гоэль М. Каушик В. и соавт. АСУ Омега. 2021 г., 20 декабря; 7(1):70-84. doi: 10.1021/acsomega.1c03216. Электронная коллекция 2022 11 января. АСУ Омега. 2021. PMID: 35036680 Бесплатная статья ЧВК.
Характеристика конъюгатов между α-лактальбумином и бензилизотиоцианатом — влияние на молекулярную структуру и протеолитическую стабильность.
Шпеттель Дж.
 , Брокельт Дж., Фальке С., Рон С.
Шпеттель Дж. и др.
Молекулы. 2021 15 октября; 26 (20): 6247. doi: 10,3390/молекулы26206247.
Молекулы. 2021.
PMID: 34684828
Бесплатная статья ЧВК.
, Брокельт Дж., Фальке С., Рон С.
Шпеттель Дж. и др.
Молекулы. 2021 15 октября; 26 (20): 6247. doi: 10,3390/молекулы26206247.
Молекулы. 2021.
PMID: 34684828
Бесплатная статья ЧВК.Эволюция складчатого ландшафта эффекторных каспаз.
Шреста С., Кларк А.С. Шреста С. и др. Дж. Биол. Хим. 2021 ноябрь; 297(5):101249. doi: 10.1016/j.jbc.2021.101249. Epub 2021 28 сентября. Дж. Биол. Хим. 2021. PMID: 34592312 Бесплатная статья ЧВК.
Рефолдинг/развертывание белка, опосредованного вихревой жидкостью, контролировалось in situ с использованием эмиссионного биозонда, индуцированного агрегацией.
Hu Q, Hu H, Zhang X, Fan K, Hong Y, Raston CL, Tang Y. Ху Кью и др. Молекулы. 2021 14 июля; 26 (14): 4273.